

Temporal asymmetries in auditory coding and perception reflect multi-layered nonlinearities
- PMID: 27580932
- PMCID: PMC5025791
- DOI: 10.1038/ncomms12682
Temporal asymmetries in auditory coding and perception reflect multi-layered nonlinearities
Abstract
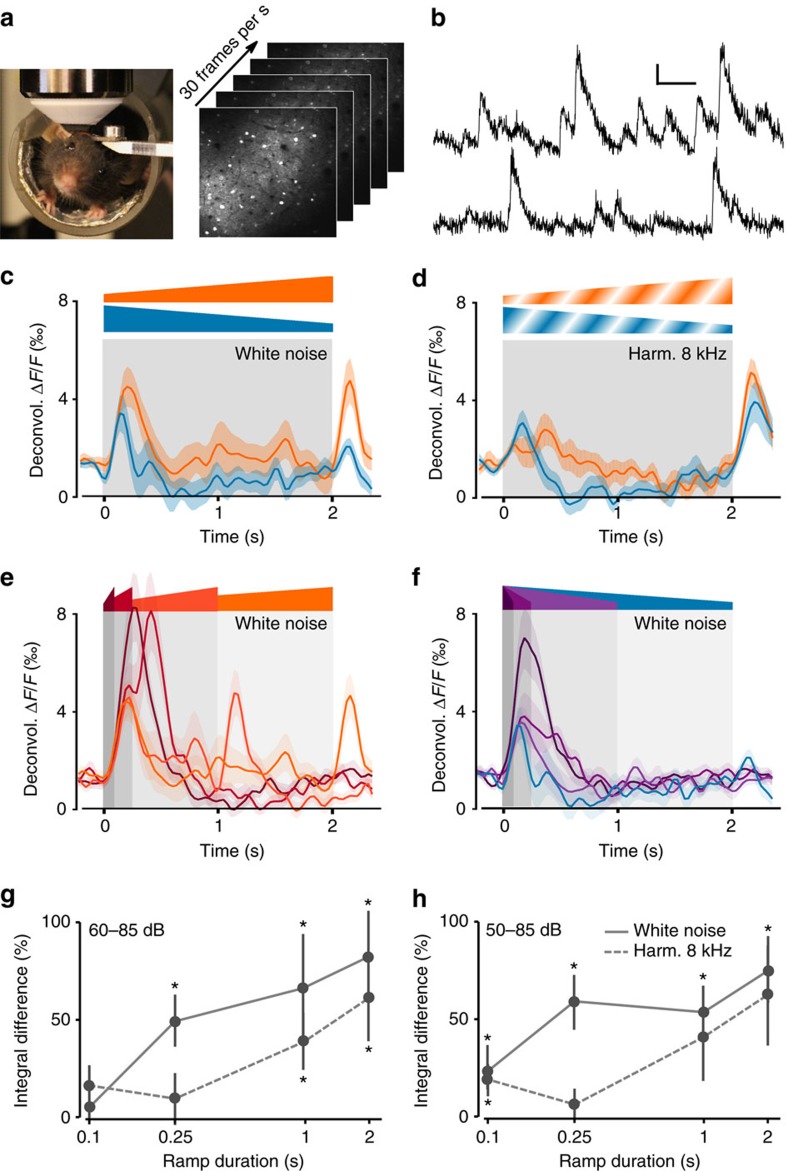
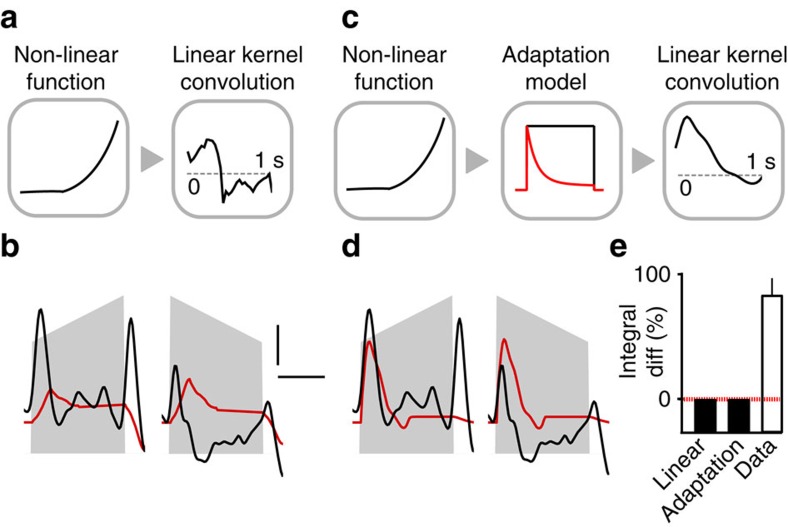
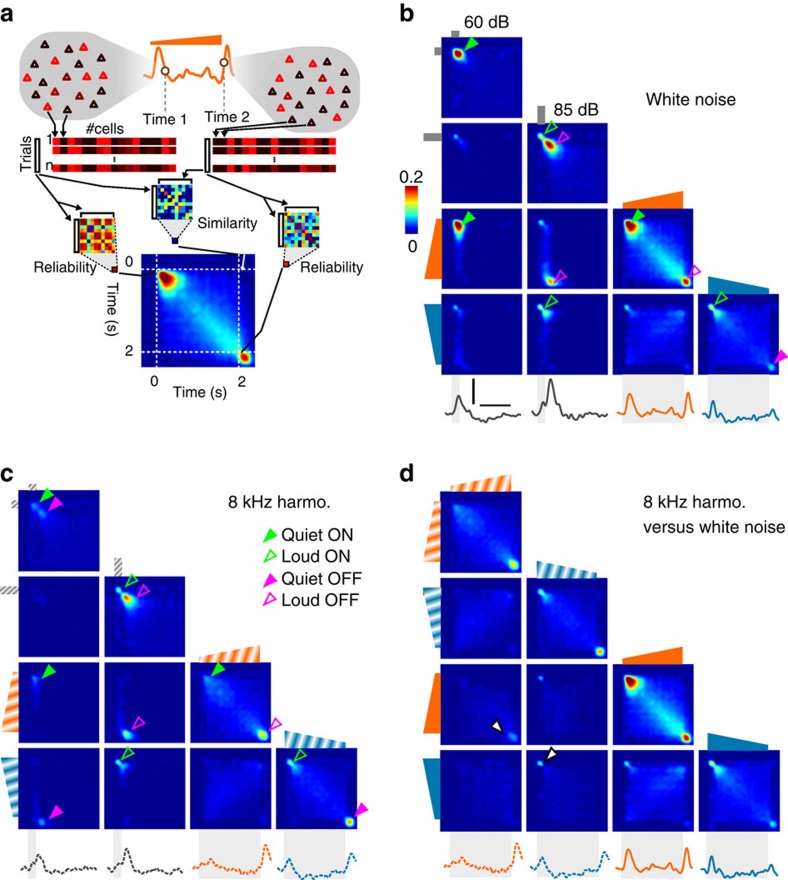
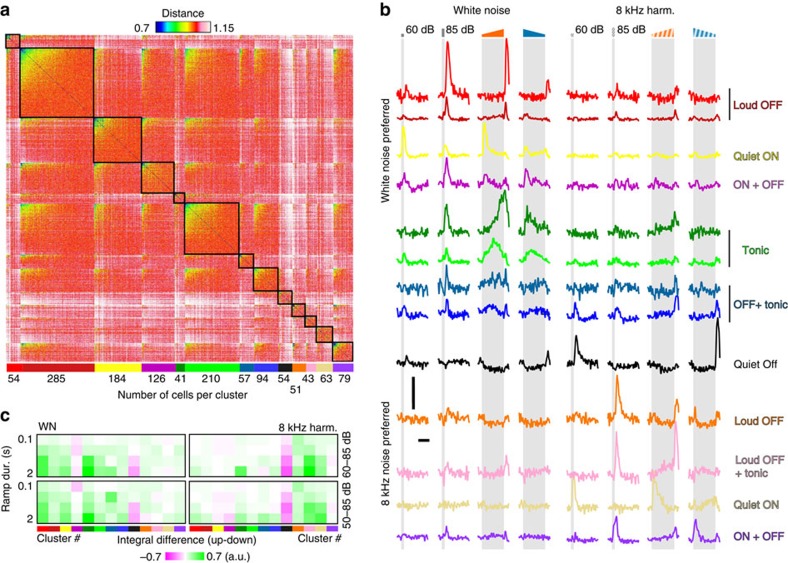
Sound recognition relies not only on spectral cues, but also on temporal cues, as demonstrated by the profound impact of time reversals on perception of common sounds. To address the coding principles underlying such auditory asymmetries, we recorded a large sample of auditory cortex neurons using two-photon calcium imaging in awake mice, while playing sounds ramping up or down in intensity. We observed clear asymmetries in cortical population responses, including stronger cortical activity for up-ramping sounds, which matches perceptual saliency assessments in mice and previous measures in humans. Analysis of cortical activity patterns revealed that auditory cortex implements a map of spatially clustered neuronal ensembles, detecting specific combinations of spectral and intensity modulation features. Comparing different models, we show that cortical responses result from multi-layered nonlinearities, which, contrary to standard receptive field models of auditory cortex function, build divergent representations of sounds with similar spectral content, but different temporal structure.
Figures
Similar articles
-
Encoding of natural sounds at multiple spectral and temporal resolutions in the human auditory cortex.PLoS Comput Biol. 2014 Jan;10(1):e1003412. doi: 10.1371/journal.pcbi.1003412. Epub 2014 Jan 2. PLoS Comput Biol. 2014. PMID: 24391486 Free PMC article.
-
How do auditory cortex neurons represent communication sounds?Hear Res. 2013 Nov;305:102-12. doi: 10.1016/j.heares.2013.03.011. Epub 2013 Apr 17. Hear Res. 2013. PMID: 23603138 Review.
-
Hearing of modulation in sounds.Physiol Rev. 1982 Jul;62(3):894-975. doi: 10.1152/physrev.1982.62.3.894. Physiol Rev. 1982. PMID: 7045902 Review. No abstract available.
-
A Hierarchy of Time Scales for Discriminating and Classifying the Temporal Shape of Sound in Three Auditory Cortical Fields.J Neurosci. 2018 Aug 1;38(31):6967-6982. doi: 10.1523/JNEUROSCI.2871-17.2018. Epub 2018 Jun 28. J Neurosci. 2018. PMID: 29954851 Free PMC article.
-
Emergence and function of cortical offset responses in sound termination detection.Elife. 2021 Dec 15;10:e72240. doi: 10.7554/eLife.72240. Elife. 2021. PMID: 34910627 Free PMC article.
Cited by
-
Phasic Off responses of auditory cortex underlie perception of sound duration.Cell Rep. 2021 Apr 20;35(3):109003. doi: 10.1016/j.celrep.2021.109003. Cell Rep. 2021. PMID: 33882311 Free PMC article.
-
Age-related changes in sound onset and offset intensity coding in auditory cortical fields A1 and CL of rhesus macaques.J Neurophysiol. 2020 Mar 1;123(3):1015-1025. doi: 10.1152/jn.00373.2019. Epub 2020 Jan 29. J Neurophysiol. 2020. PMID: 31995426 Free PMC article.
-
Spontaneous Mouse Behavior in Presence of Dissonance and Acoustic Roughness.Front Behav Neurosci. 2020 Oct 8;14:588834. doi: 10.3389/fnbeh.2020.588834. eCollection 2020. Front Behav Neurosci. 2020. PMID: 33132864 Free PMC article.
-
A generic deviance detection principle for cortical On/Off responses, omission response, and mismatch negativity.Biol Cybern. 2019 Dec;113(5-6):475-494. doi: 10.1007/s00422-019-00804-x. Epub 2019 Aug 19. Biol Cybern. 2019. PMID: 31428855 Free PMC article.
-
Neural Mechanisms Underlying the Auditory Looming Bias.Audit Percept Cogn. 2021 Apr 3;4(1-2):60-73. doi: 10.1080/25742442.2021.1977582. Epub 2021 Sep 20. Audit Percept Cogn. 2021. PMID: 35494218 Free PMC article.
References
-
- Helmholtz H. v. & Ellis A. J. On the Sensations of Tone as a Physiological Basis for the Theory of Music 2nd edn Longmans, Green (1885).
-
- Lewis J. W. et al.. Human brain regions involved in recognizing environmental sounds. Cereb. Cortex. 14, 1008–1021 (2004). - PubMed
-
- McBeath M. K. & Neuhoff J. G. The Doppler effect is not what you think it is: dramatic pitch change due to dynamic intensity change. Psychon. Bull. Rev. 9, 306–313 (2002). - PubMed
-
- Nelken I., Rotman Y. & Bar Yosef O. Responses of auditory-cortex neurons to structural features of natural sounds. Nature 397, 154–157 (1999). - PubMed
-
- Theunissen F. E. & Elie J. E. Neural processing of natural sounds. Nat. Rev. Neurosci. 15, 355–366 (2014). - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
