

Human adaptation and evolution by segmental duplication
- PMID: 27584858
- PMCID: PMC5161654
- DOI: 10.1016/j.gde.2016.08.001
Human adaptation and evolution by segmental duplication
Abstract
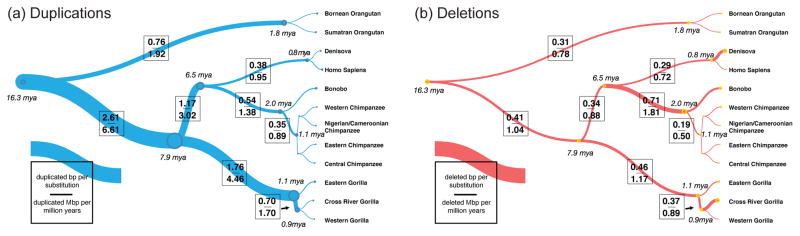
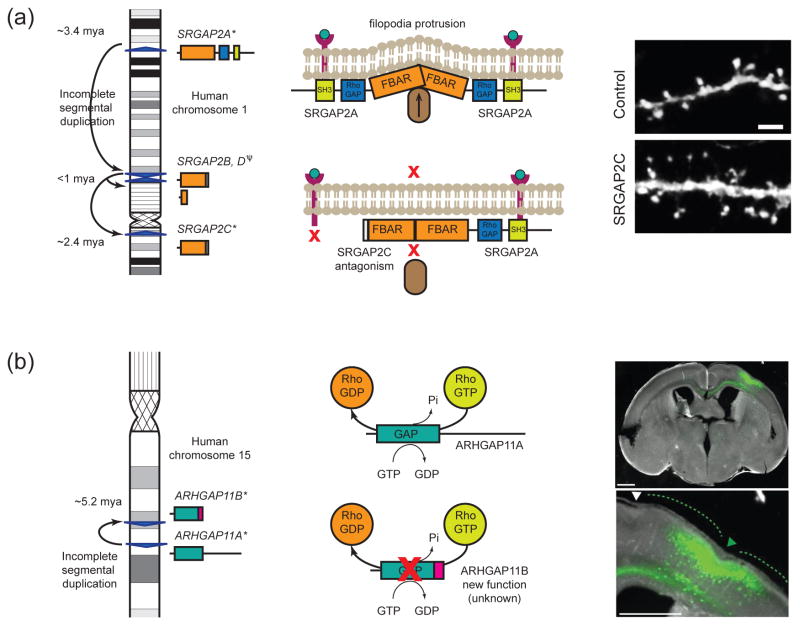
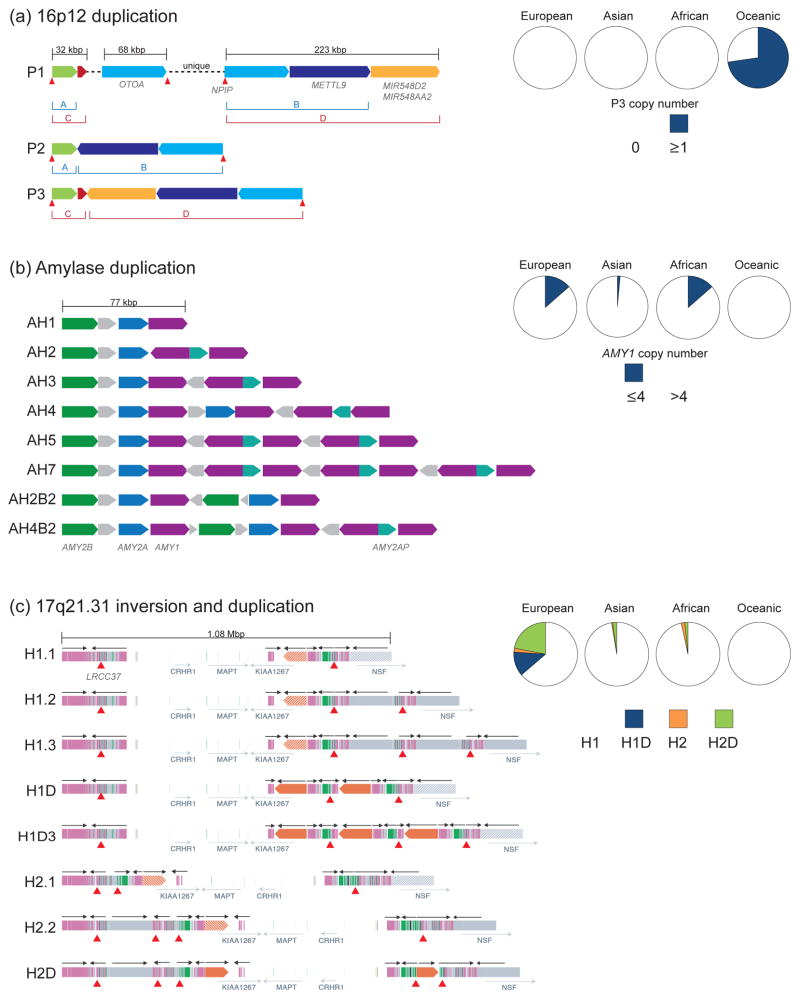
Duplications are the primary force by which new gene functions arise and provide a substrate for large-scale structural variation. Analysis of thousands of genomes shows that humans and great apes have more genetic differences in content and structure over recent segmental duplications than any other euchromatic region. Novel human-specific duplicated genes, ARHGAP11B and SRGAP2C, have recently been described with a potential role in neocortical expansion and increased neuronal spine density. Large segmental duplications and the structural variants they promote are also frequently stratified between human populations with a subset being subjected to positive selection. The impact of recent duplications on human evolution and adaptation is only beginning to be realized as new technologies enhance their discovery and accurate genotyping.
Copyright © 2016 Elsevier Ltd. All rights reserved.
Figures
References
-
- Haldane JBS. The causes of evolution. London, New York etc: Longmans, Green and co; 1932.
-
- Ohno S. Evolution by gene duplication. London, New York: Allen & Unwin; Springer-Verlag; 1970.
-
- Lynch M, Conery JS. The evolutionary fate and consequences of duplicate genes. Science. 2000;290:1151–1155. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
