

Gata3 restrains B cell proliferation and cooperates with p18INK4c to repress B cell lymphomagenesis
- PMID: 27588406
- PMCID: PMC5325421
- DOI: 10.18632/oncotarget.11746
Gata3 restrains B cell proliferation and cooperates with p18INK4c to repress B cell lymphomagenesis
Abstract
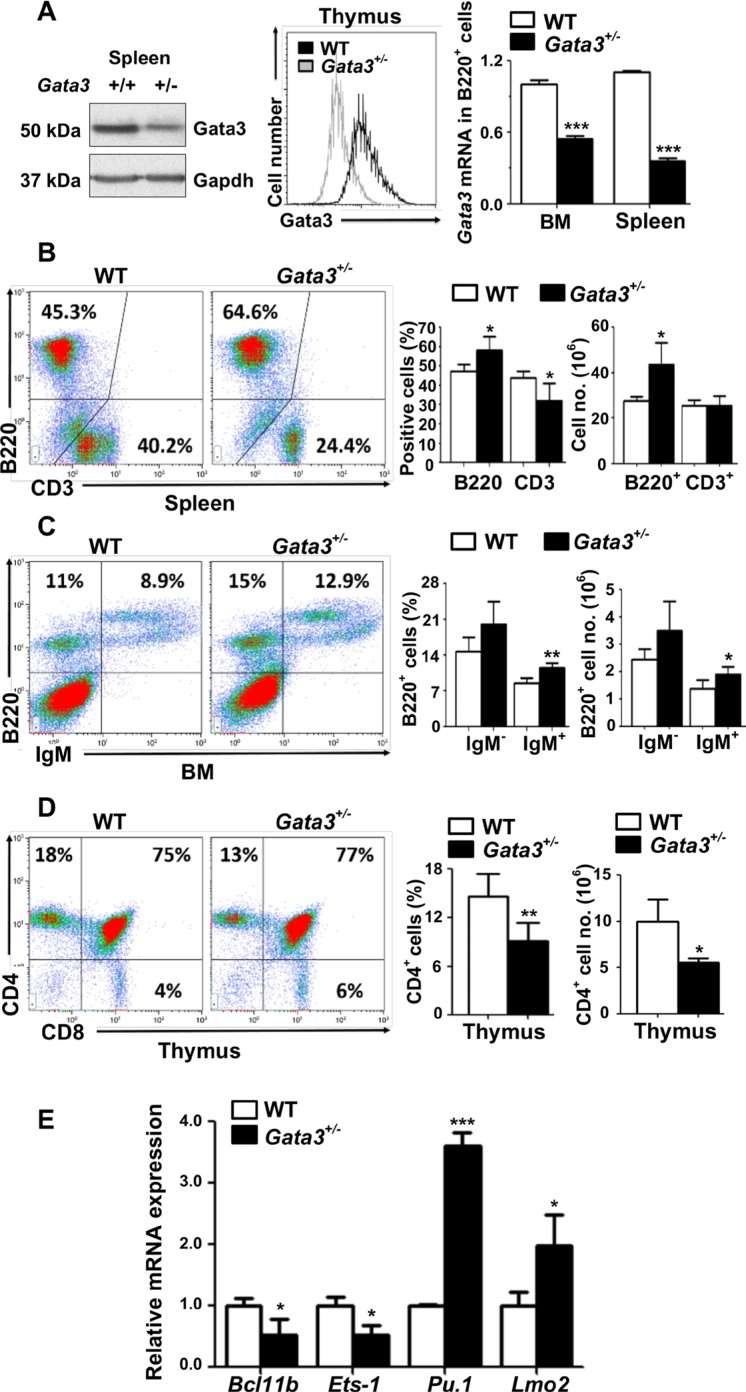
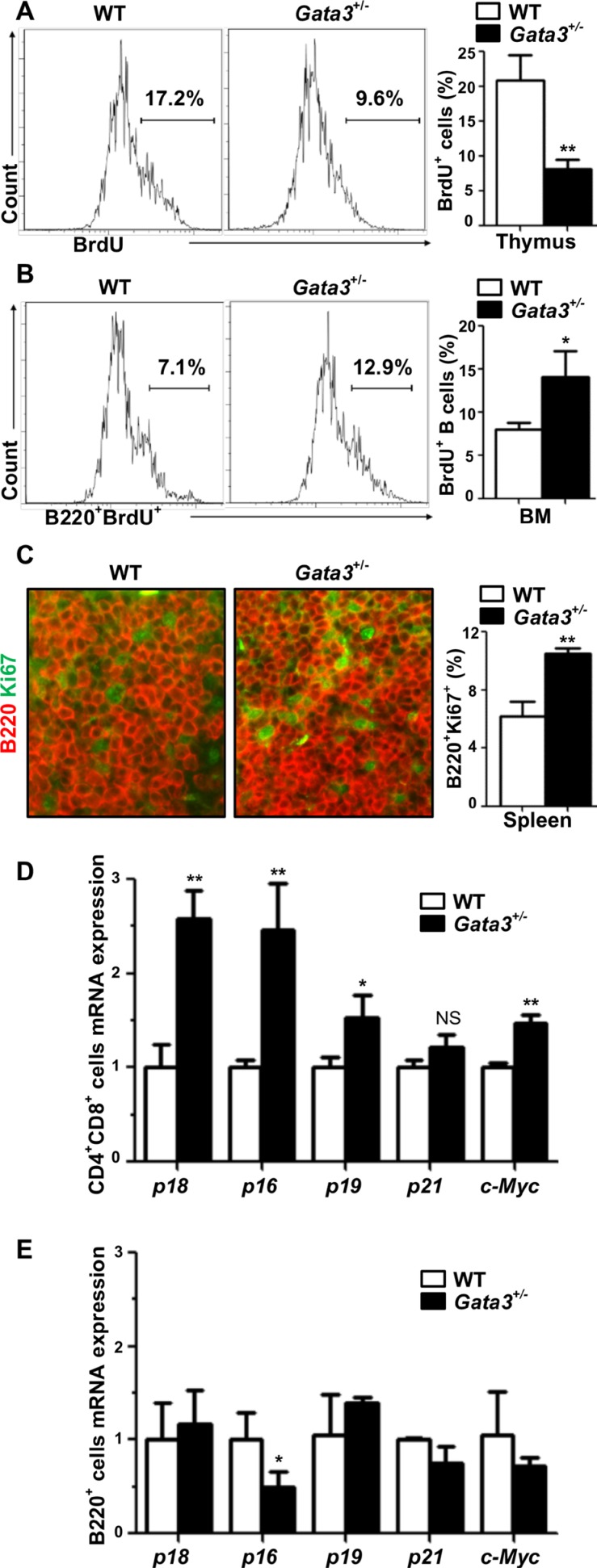
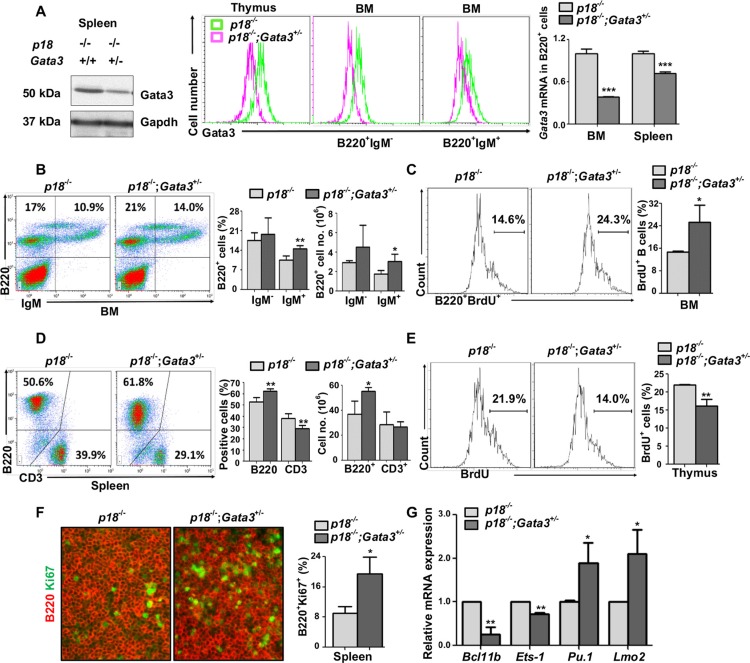
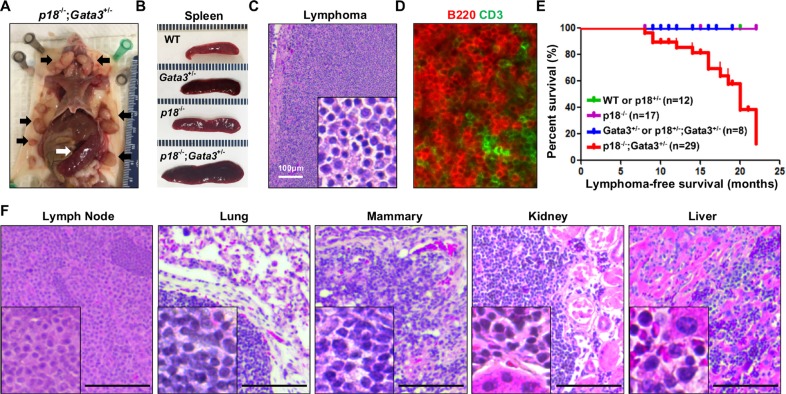
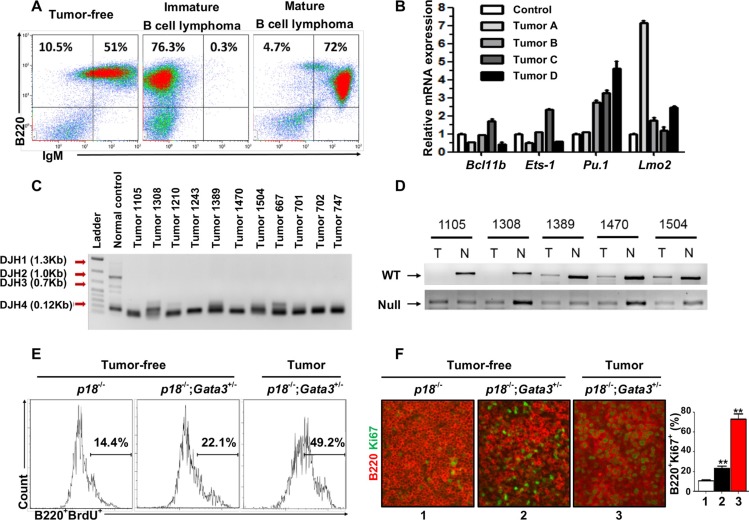
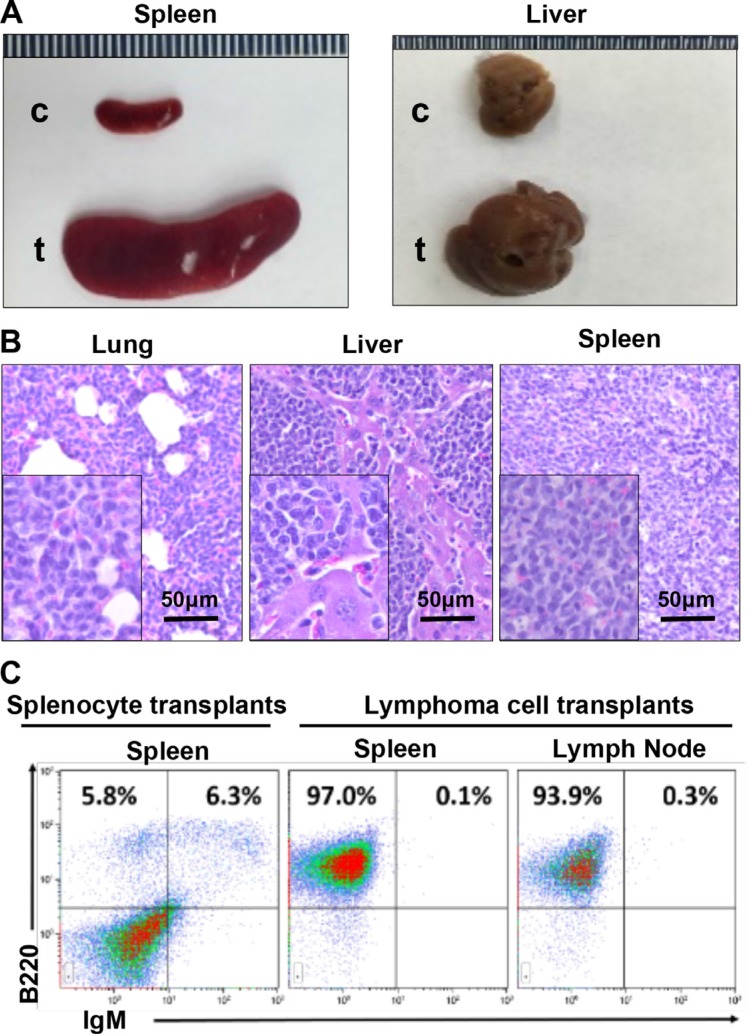
GATA3, a lineage specifier, controls lymphoid cell differentiation and its function in T cell commitment and development has been extensively studied. GATA3 promotes T cell specification by repressing B cell potential in pro T cells and decreased GATA3 expression is essential for early B cell commitment. Inherited genetic variation in GATA3 has been associated with lymphoma susceptibility. However, it remains elusive how the loss of function of GATA3 promotes B cell development and induces B cell lymphomas. In this study, we found that haploid loss of Gata3 by heterozygous germline deletion increased B cell populations in the bone marrow (BM) and spleen, and decreased CD4 T cell populations in the thymus, confirming that Gata3 promotes T and suppresses B cell development. We discovered that haploid loss of Gata3 reduced thymocyte proliferation with induction of p18Ink4c (p18), an inhibitor of CDK4 and CDK6, but enhanced B cell proliferation in the BM and spleen independent of p18. Loss of p18 partially restored Gata3 deficient thymocyte proliferation, but further stimulated Gata3 deficient B cell proliferation in the BM and spleen. Furthermore, we discovered that haploid loss of Gata3 in p18 deficient mice led to the development of B cell lymphomas that were capable of rapidly regenerating tumors when transplanted into immunocompromised mice. These results indicate that Gata3 deficiency promotes B cell differentiation and proliferation, and cooperates with p18 loss to induce B cell lymphomas. This study, for the first time, reveals that Gata3 is a tumor suppressor specifically in B cell lymphomagenesis.
Keywords: B cell; Gata3; lymphoma; p18INK4c.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
References
-
- Horner MJ RL, Krapcho M, Neyman N, Aminou R, Howlader N, Altekruse SF, Feuer EJ, Huang L, Mariotto A, Miller BA, Lewis DR, Eisner MP, Stinchcomb DG, et al. SEER Cancer Statistics Review 1975–2006. National Cancer Institute; Bethesda MD: based on November 2008 SEER data submission posted to the SEER web site 2009.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
