

Global Mapping of Small RNA-Target Interactions in Bacteria
- PMID: 27588604
- PMCID: PMC5145812
- DOI: 10.1016/j.molcel.2016.07.026
Global Mapping of Small RNA-Target Interactions in Bacteria
Abstract
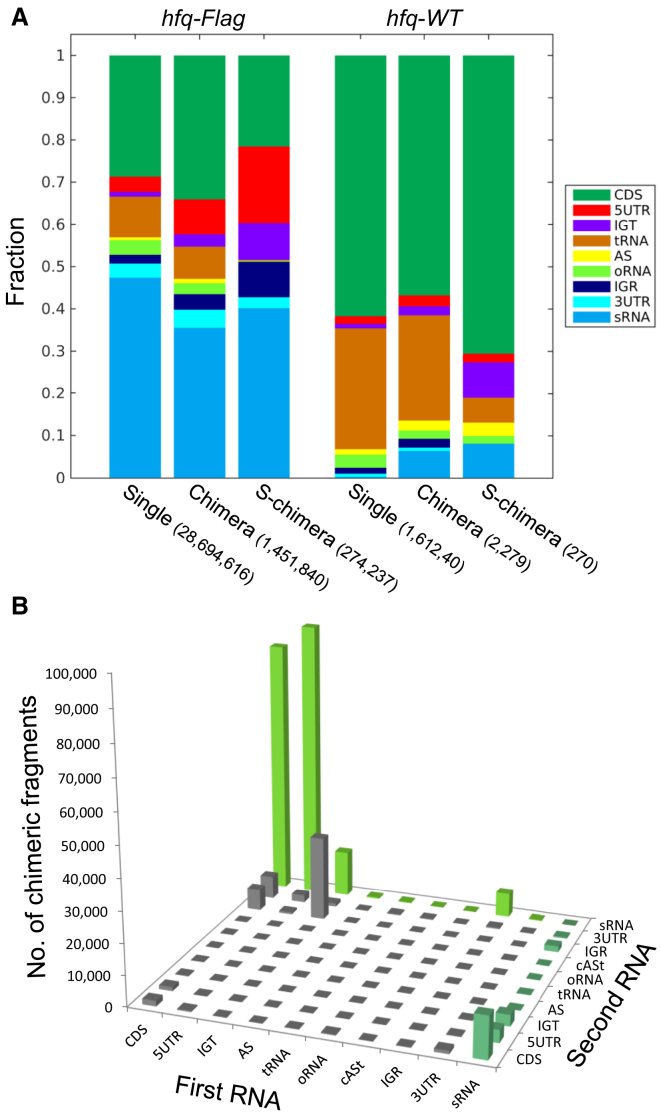
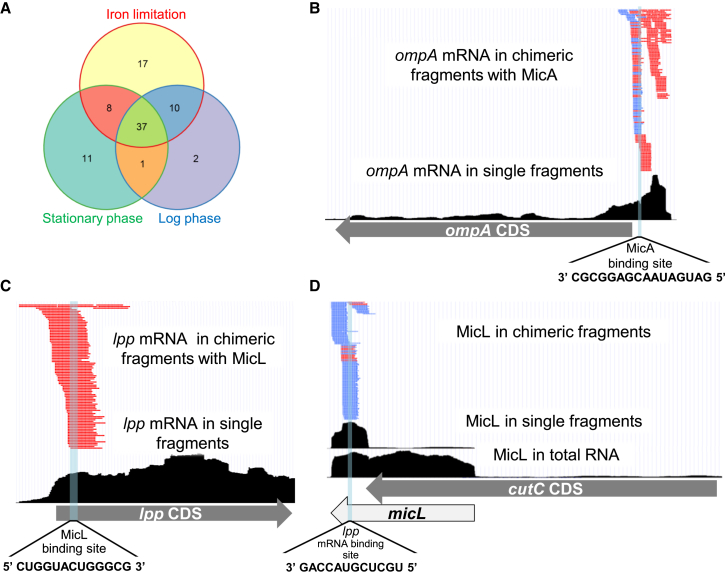
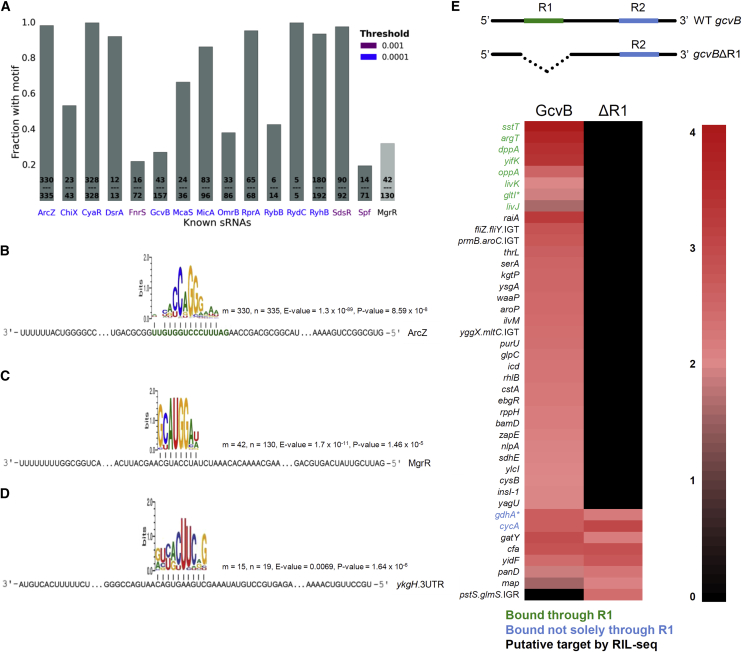
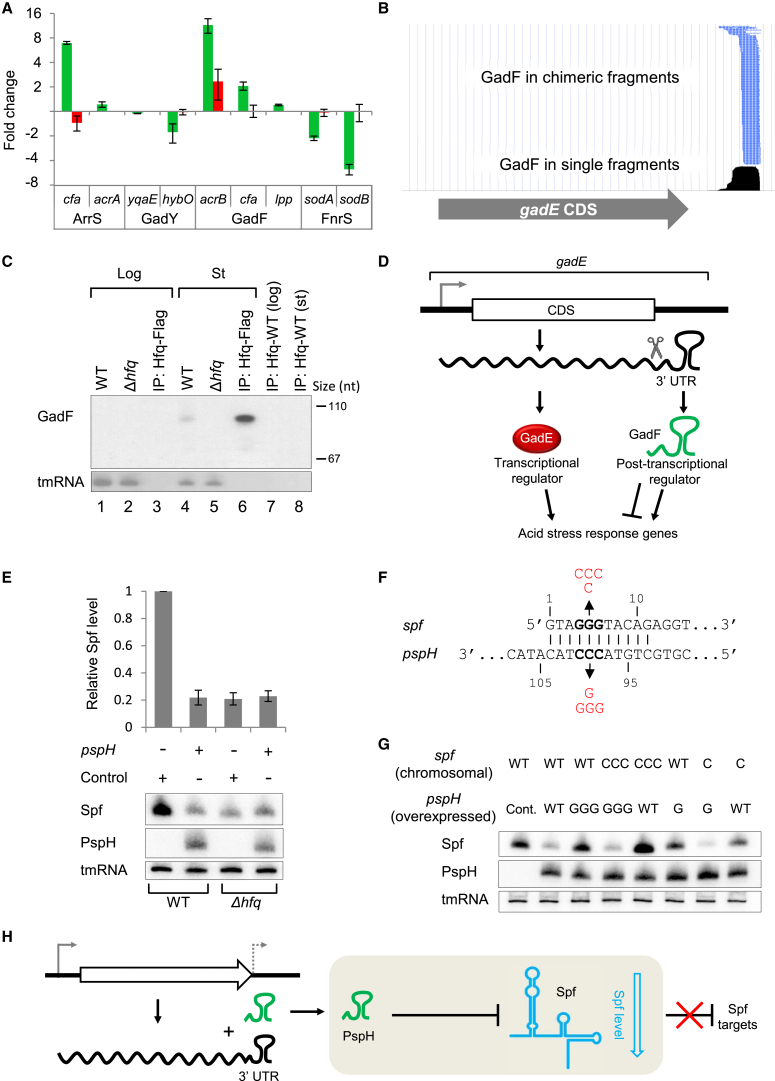
Small RNAs (sRNAs) associated with the RNA chaperon protein Hfq are key posttranscriptional regulators of gene expression in bacteria. Deciphering the sRNA-target interactome is an essential step toward understanding the roles of sRNAs in the cellular networks. We developed a broadly applicable methodology termed RIL-seq (RNA interaction by ligation and sequencing), which integrates experimental and computational tools for in vivo transcriptome-wide identification of interactions involving Hfq-associated sRNAs. By applying this methodology to Escherichia coli we discovered an extensive network of interactions involving RNA pairs showing sequence complementarity. We expand the ensemble of targets for known sRNAs, uncover additional Hfq-bound sRNAs encoded in various genomic regions along with their trans encoded targets, and provide insights into binding and possible cycling of RNAs on Hfq. Comparison of the sRNA interactome under various conditions has revealed changes in the sRNA repertoire as well as substantial re-wiring of the network between conditions.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures




Comment in
-
Global snapshots of bacterial RNA networks.EMBO J. 2017 Feb 1;36(3):245-247. doi: 10.15252/embj.201696072. Epub 2016 Dec 28. EMBO J. 2017. PMID: 28031253 Free PMC article.
References
-
- Argaman L., Hershberg R., Vogel J., Bejerano G., Wagner E.G., Margalit H., Altuvia S. Novel small RNA-encoding genes in the intergenic regions of Escherichia coli. Curr. Biol. 2001;11:941–950. - PubMed
-
- Bailey T.L., Gribskov M. Methods and statistics for combining motif match scores. J. Comput. Biol. 1998;5:211–221. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
