

Thalamic Inhibition: Diverse Sources, Diverse Scales
- PMID: 27589879
- PMCID: PMC5048590
- DOI: 10.1016/j.tins.2016.08.001
Thalamic Inhibition: Diverse Sources, Diverse Scales
Abstract
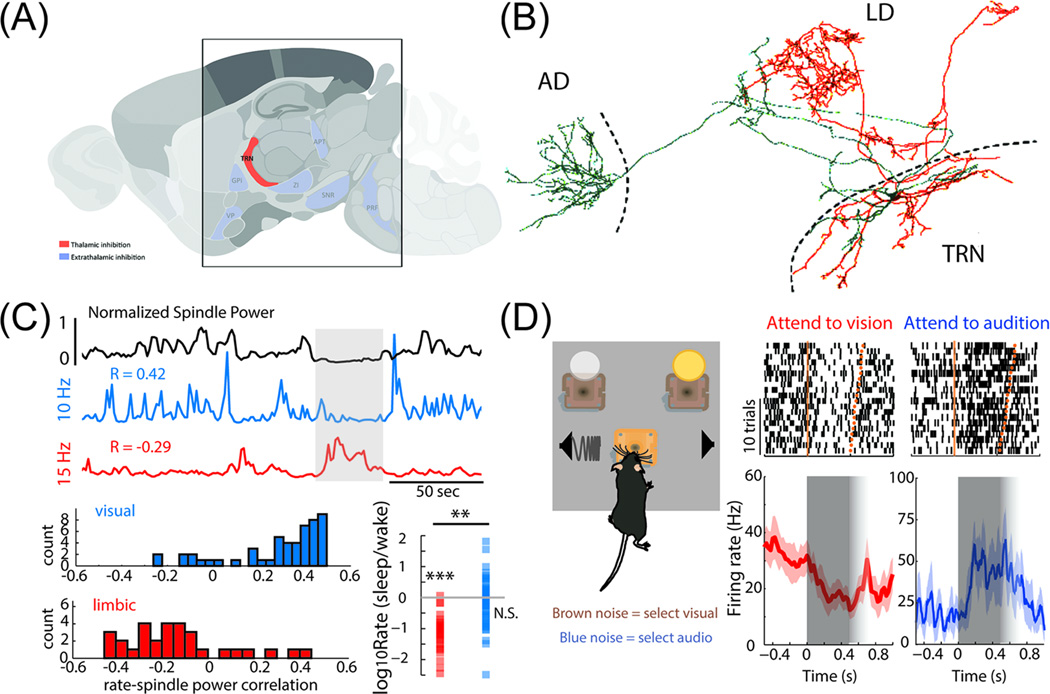
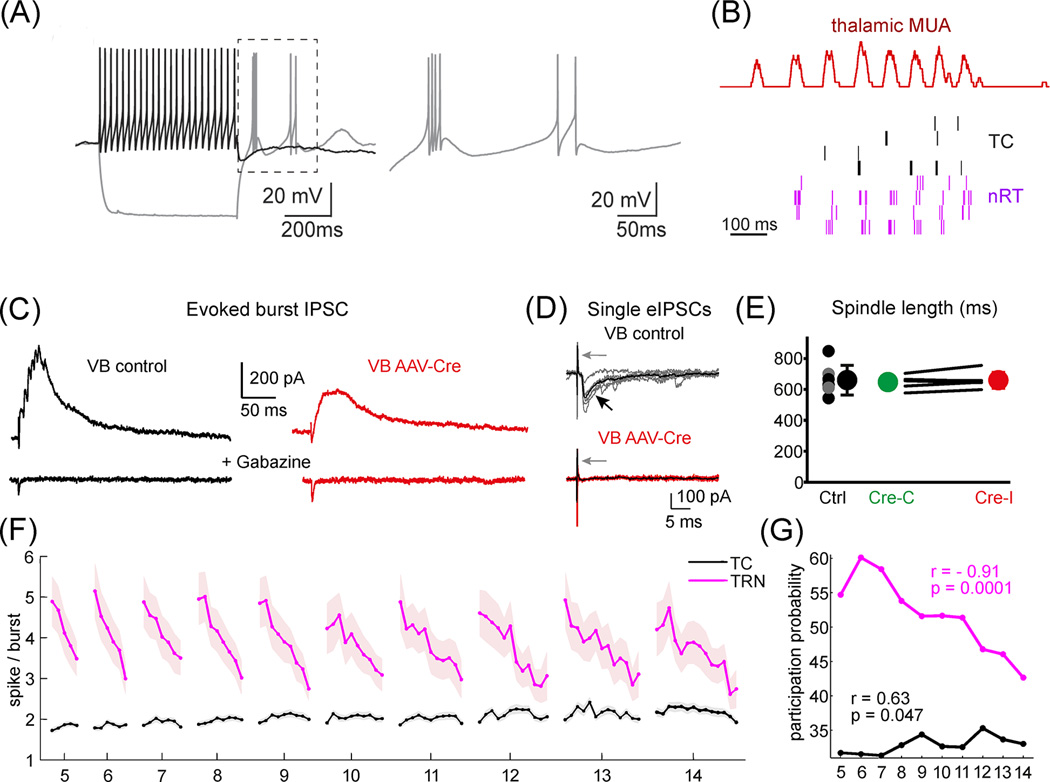
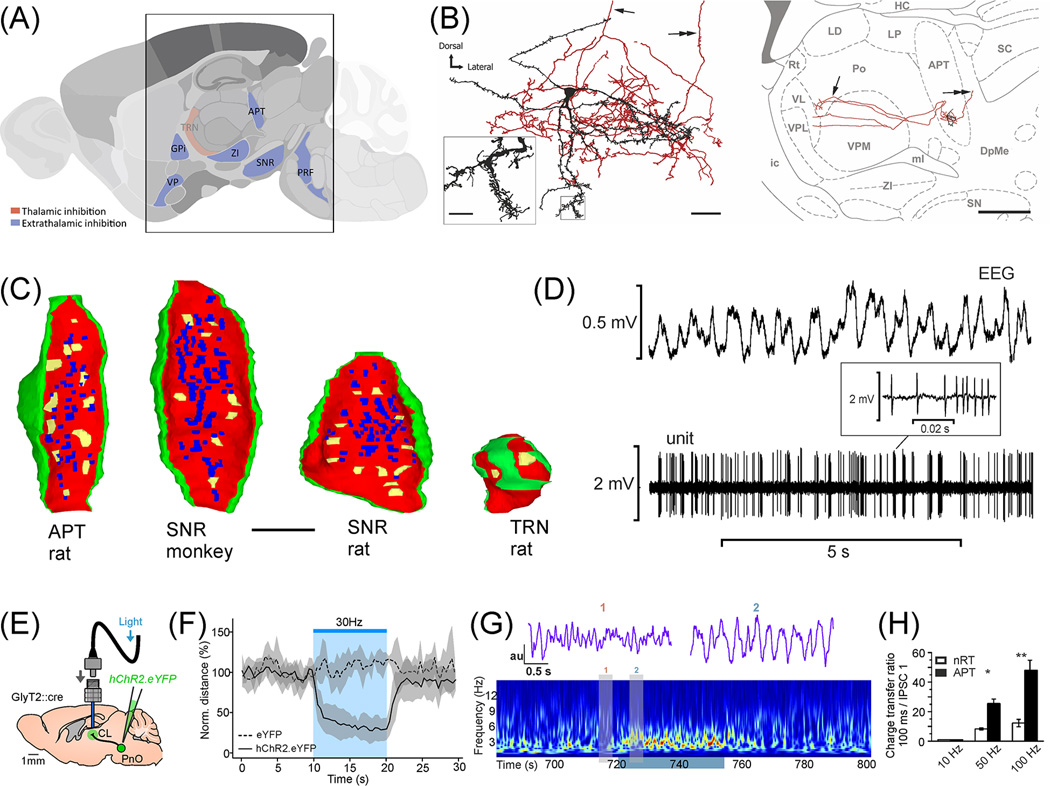
The thalamus is the major source of cortical inputs shaping sensation, action, and cognition. Thalamic circuits are targeted by two major inhibitory systems: the thalamic reticular nucleus (TRN) and extrathalamic inhibitory (ETI) inputs. A unifying framework of how these systems operate is currently lacking. Here, we propose that TRN circuits are specialized to exert thalamic control at different spatiotemporal scales. Local inhibition of thalamic spike rates prevails during attentional selection, whereas global inhibition more likely prevails during sleep. In contrast, the ETI (arising from basal ganglia, zona incerta (ZI), anterior pretectum, and pontine reticular formation) provides temporally precise and focal inhibition, impacting spike timing. Together, these inhibitory systems allow graded control of thalamic output, enabling thalamocortical operations to dynamically match ongoing behavioral demands.
Keywords: GABA; basal ganglia; reticular thalamic nucleus; thalamocortical.
Copyright © 2016 Elsevier Ltd. All rights reserved.
Figures
References
-
- Ketz NA, et al. Thalamic pathways underlying prefrontal cortex-medial temporal lobe oscillatory interactions. Trends Neurosci. 2015;38:3–12. - PubMed
-
- Schiff ND, et al. Behavioural improvements with thalamic stimulation after severe traumatic brain injury. Nature. 2007;448:600–603. - PubMed
-
- Hirsch JA, et al. How inhibitory circuits in the thalamus serve vision. Annu. Rev. Neurosci. 2015;38:309–329. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
