

Role of the Conserved Disulfide Bridge in Class A Carbapenemases
- PMID: 27590339
- PMCID: PMC5063999
- DOI: 10.1074/jbc.M116.749648
Role of the Conserved Disulfide Bridge in Class A Carbapenemases
Abstract
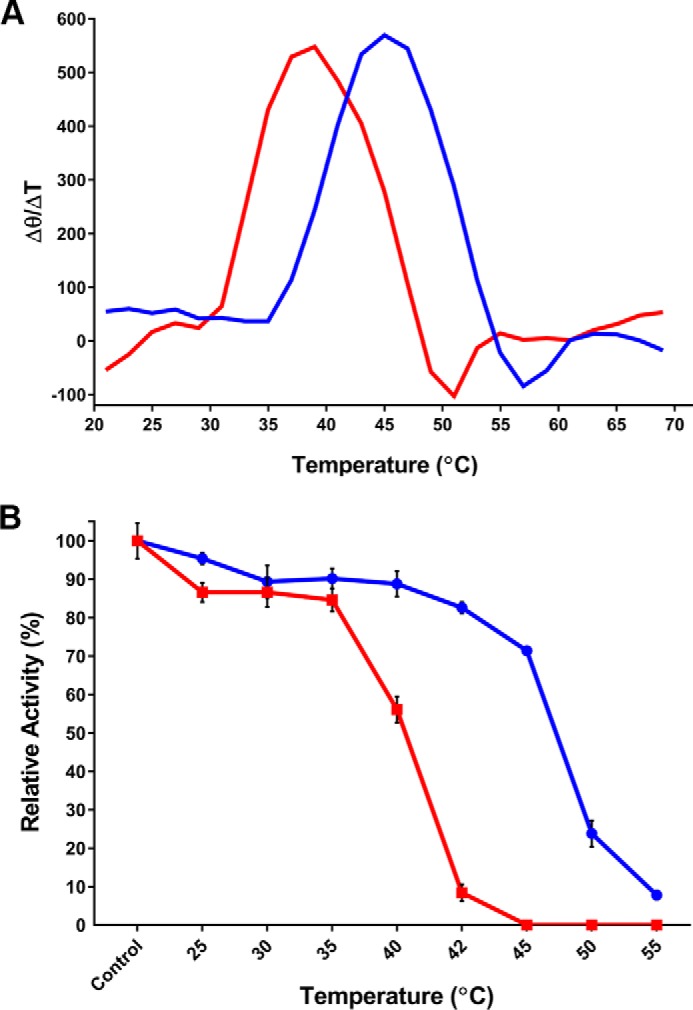
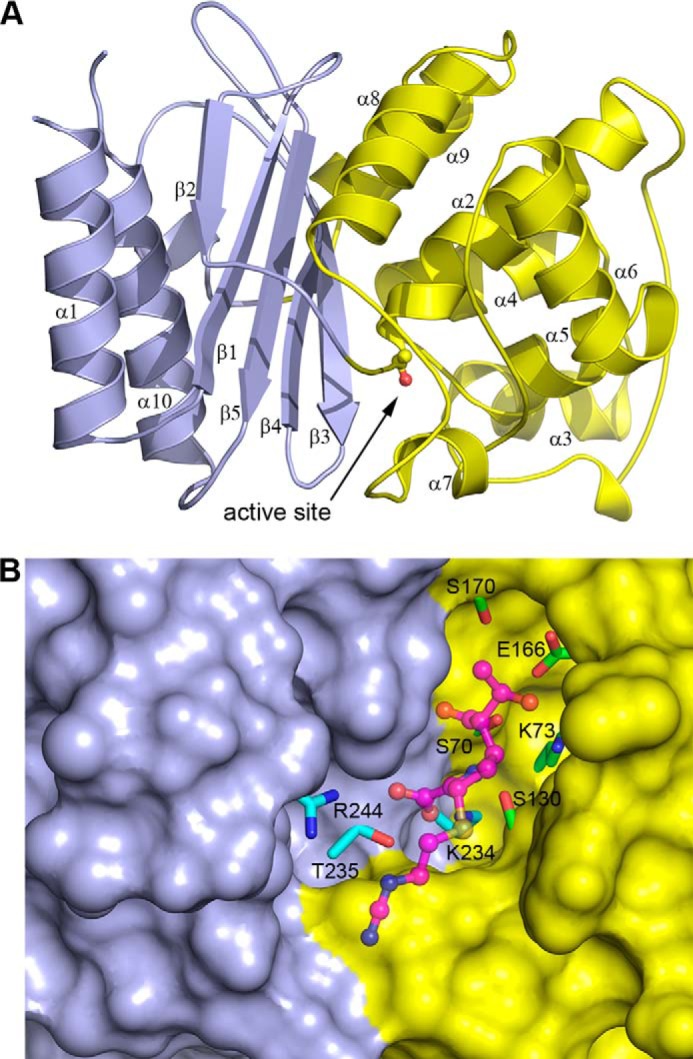
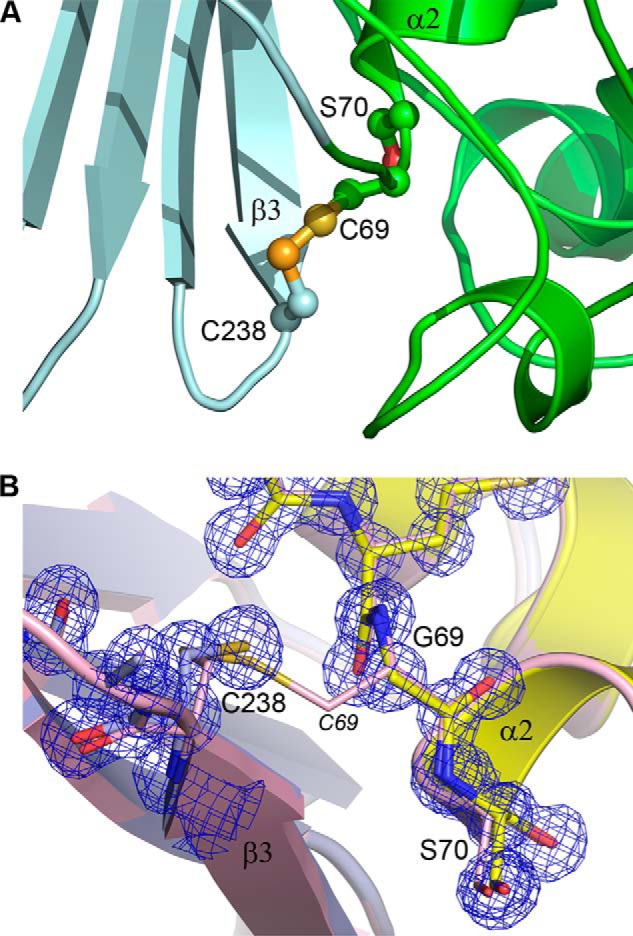
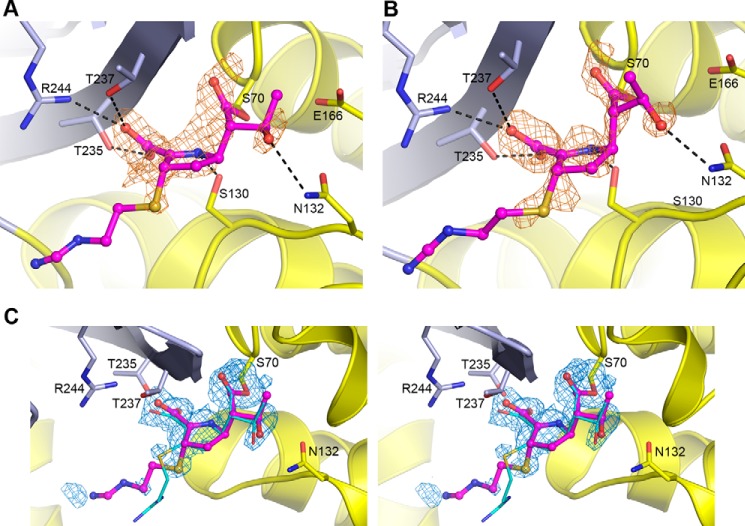
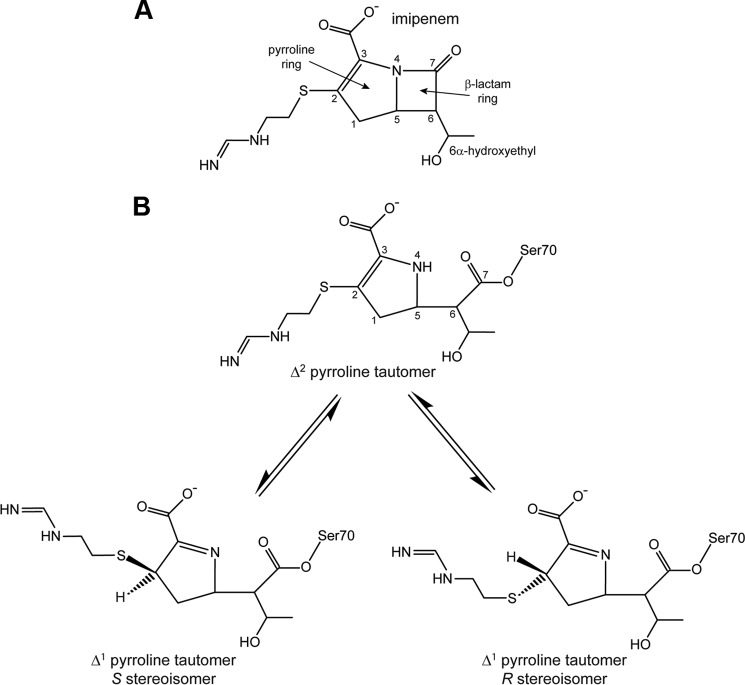
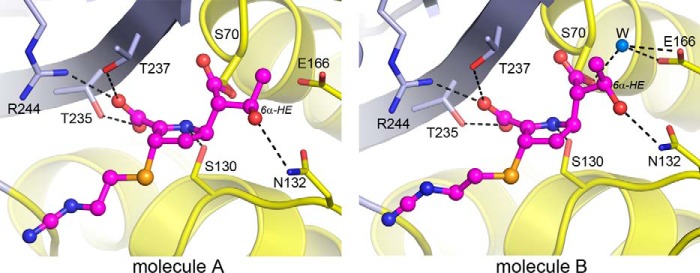
Some members of the class A β-lactamase family are capable of conferring resistance to the last resort antibiotics, carbapenems. A unique structural feature of these clinically important enzymes, collectively referred to as class A carbapenemases, is a disulfide bridge between invariant Cys69 and Cys238 residues. It was proposed that this conserved disulfide bridge is responsible for their carbapenemase activity, but this has not yet been validated. Here we show that disruption of the disulfide bridge in the GES-5 carbapenemase by the C69G substitution results in only minor decreases in the conferred levels of resistance to the carbapenem imipenem and other β-lactams. Kinetic and circular dichroism experiments with C69G-GES-5 demonstrate that this small drop in antibiotic resistance is due to a decline in the enzyme activity caused by a marginal loss of its thermal stability. The atomic resolution crystal structure of C69G-GES-5 shows that two domains of this disulfide bridge-deficient enzyme are held together by an intensive hydrogen-bonding network. As a result, the protein architecture and imipenem binding mode remain unchanged. In contrast, the corresponding hydrogen-bonding networks in NMCA, SFC-1, and SME-1 carbapenemases are less intensive, and as a consequence, disruption of the disulfide bridge in these enzymes destabilizes them, which causes arrest of bacterial growth. Our results demonstrate that the disulfide bridge is essential for stability but does not play a direct role in the carbapenemase activity of the GES family of β-lactamases. This would likely apply to all other class A carbapenemases given the high degree of their structural similarity.
Keywords: antibiotic resistance; carbapenemase; crystal structure; disulfide; enzyme kinetics; enzyme stability; hydrogen bond.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
