

A Genome-wide CRISPR Screen in Toxoplasma Identifies Essential Apicomplexan Genes
- PMID: 27594426
- PMCID: PMC5017925
- DOI: 10.1016/j.cell.2016.08.019
A Genome-wide CRISPR Screen in Toxoplasma Identifies Essential Apicomplexan Genes
Abstract
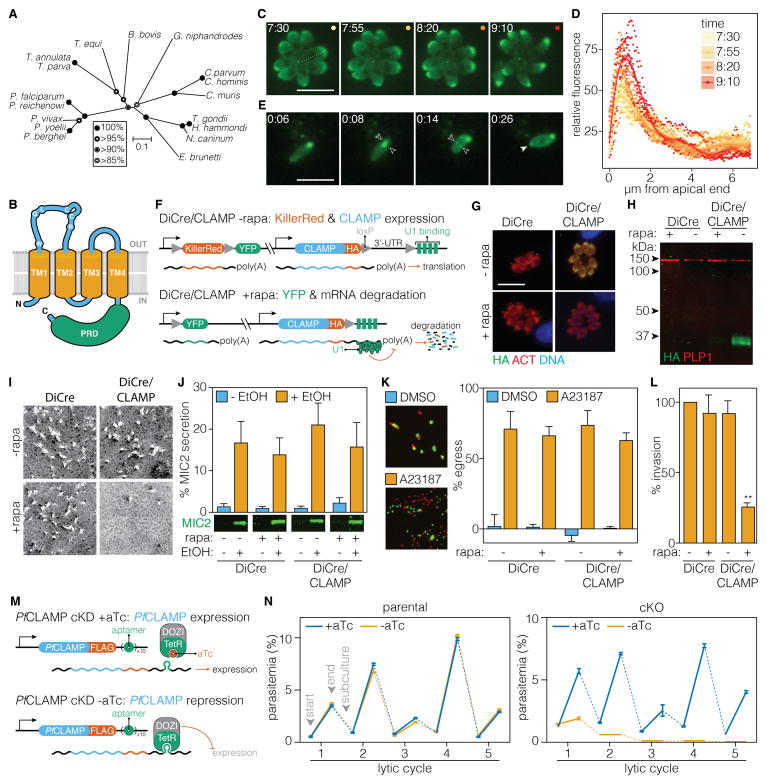
Apicomplexan parasites are leading causes of human and livestock diseases such as malaria and toxoplasmosis, yet most of their genes remain uncharacterized. Here, we present the first genome-wide genetic screen of an apicomplexan. We adapted CRISPR/Cas9 to assess the contribution of each gene from the parasite Toxoplasma gondii during infection of human fibroblasts. Our analysis defines ∼200 previously uncharacterized, fitness-conferring genes unique to the phylum, from which 16 were investigated, revealing essential functions during infection of human cells. Secondary screens identify as an invasion factor the claudin-like apicomplexan microneme protein (CLAMP), which resembles mammalian tight-junction proteins and localizes to secretory organelles, making it critical to the initiation of infection. CLAMP is present throughout sequenced apicomplexan genomes and is essential during the asexual stages of the malaria parasite Plasmodium falciparum. These results provide broad-based functional information on T. gondii genes and will facilitate future approaches to expand the horizon of antiparasitic interventions.
Keywords: Apicomplexan parasites; eukaryotic pathogen; genome-wide CRISPR screen; host-cell invasion; host-pathogen interactions; malaria; toxoplasmosis.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
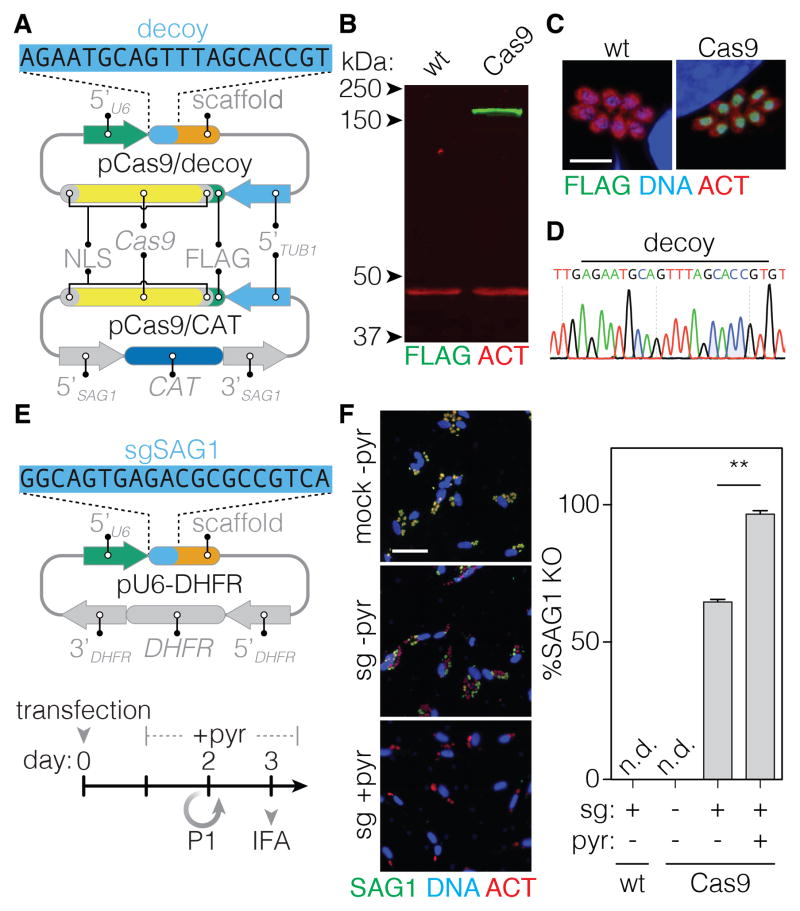

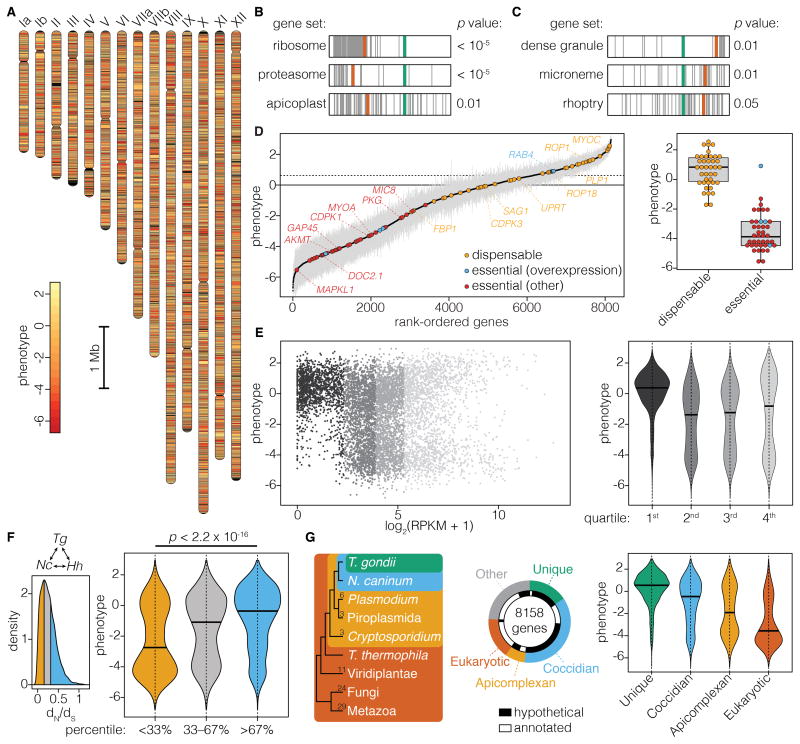
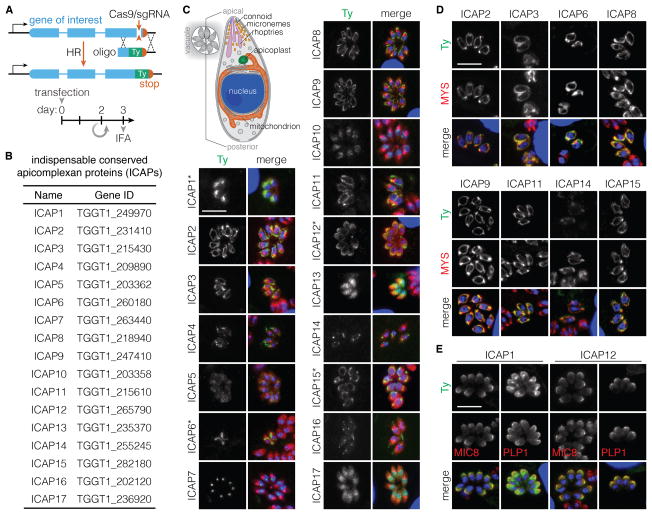
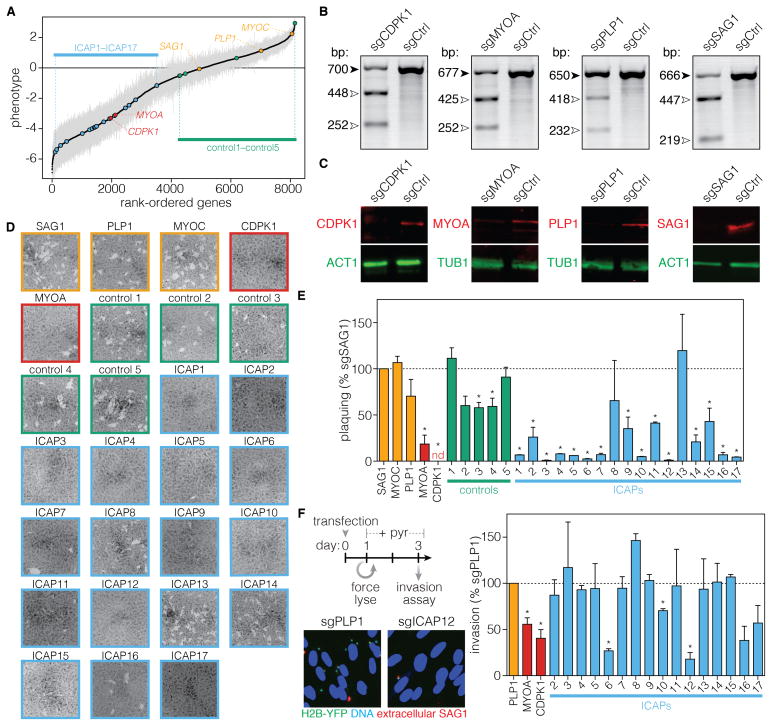
Comment in
-
mSphere of Influence: Systemic decoding gene function in Toxoplasma gondii pathogenesis-CRISPR screens and beyond.mSphere. 2023 Aug 24;8(4):e0027123. doi: 10.1128/msphere.00271-23. Epub 2023 Jun 29. mSphere. 2023. PMID: 37382434 Free PMC article.
References
-
- Achbarou A, Mercereau-Puijalon O, Autheman JM, Fortier B, Camus D, Dubremetz JF. Characterization of microneme proteins of Toxoplasma gondii. Mol Biochem Parasitol. 1991;47:223–233. - PubMed
-
- Bastin P, Bagherzadeh Z, Matthews KR, Gull K. A novel epitope tag system to study protein targeting and organelle biogenesis in Trypanosoma brucei. Mol Biochem Parasitol. 1996;77:235–239. - PubMed
-
- Bullen HE, Jia Y, Yamaryo-Botté Y, Bisio H, Zhang O, Jemelin NK, Marq JB, Carruthers V, Botté CY, Soldati-Favre D. Phosphatidic Acid-Mediated Signaling Regulates Microneme Secretion in Toxoplasma. Cell Host Microbe. 2016;19:349–360. - PubMed
-
- Burg JL, Perelman D, Kasper LH, Ware PL, Boothroyd JC. Molecular analysis of the gene encoding the major surface antigen of Toxoplasma gondii. J Immunol. 1988;141:3584–3591. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
