

CD44 and RHAMM are essential for rapid growth of bladder cancer driven by loss of Glycogen Debranching Enzyme (AGL)
- PMID: 27595989
- PMCID: PMC5011830
- DOI: 10.1186/s12885-016-2756-5
CD44 and RHAMM are essential for rapid growth of bladder cancer driven by loss of Glycogen Debranching Enzyme (AGL)
Abstract
Background: Loss of Amylo-alpha-1-6-glucosidase-4-alpha-glucanotransferase (AGL) drives rapid proliferation of bladder cancer cells by upregulating Hyaluronic acid(HA) Synthase (HAS2) mediated HA synthesis. However the role of HA receptors CD44 and Hyaluronan Mediated Motility Receptor (RHAMM) in regulating the growth of bladder cancer cells driven by loss of AGL has not been studied.
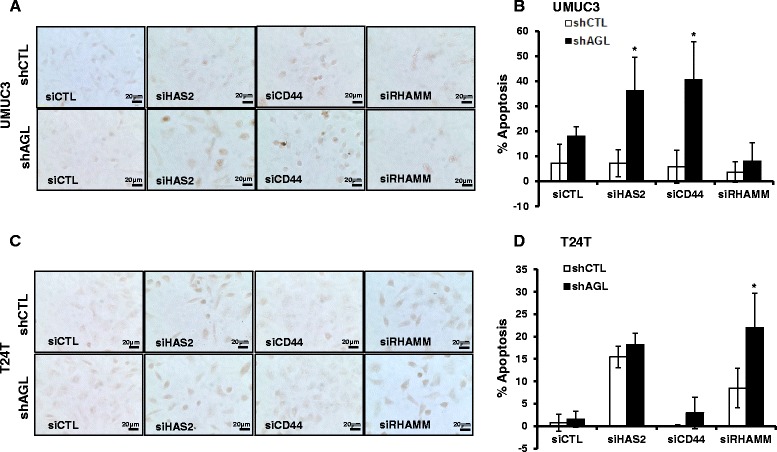
Methods: Western blot analysis and Terminal deoxynucleotidyl transferase (TdT) dUTP Nick-End Labeling (TUNEL) assay was carried out to study cellular apoptosis with HAS2, CD44 and RHAMM loss in bladder cancer cells with and without AGL expression. Proliferation and softagar assays were carried out to study cellular anchorage dependent and independent growth. Clinicopathologic analysis was carried out on bladder cancer patient datasets.
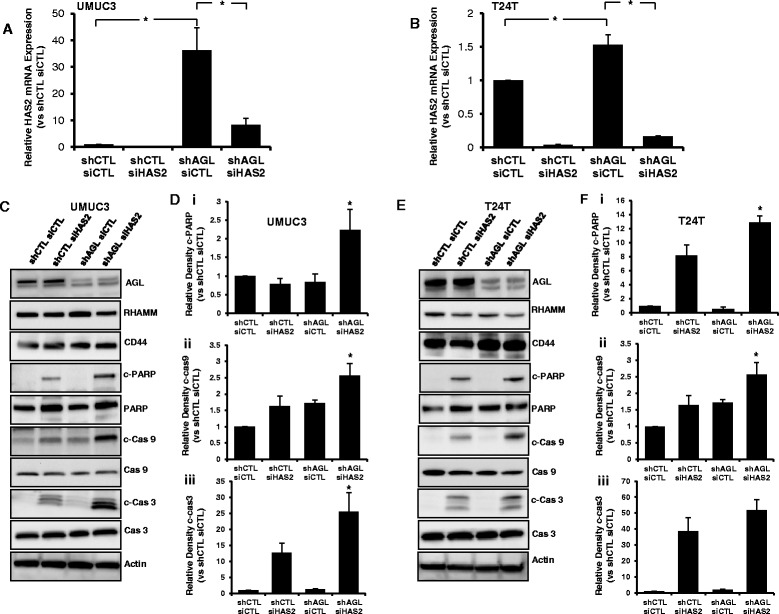
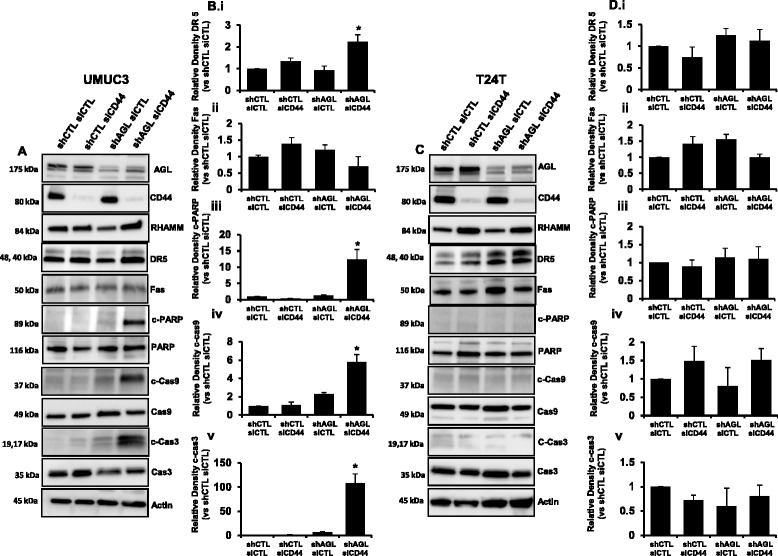
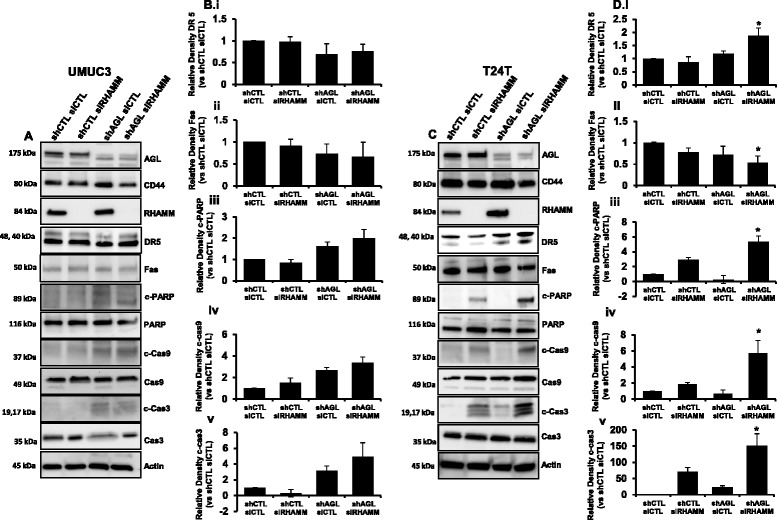
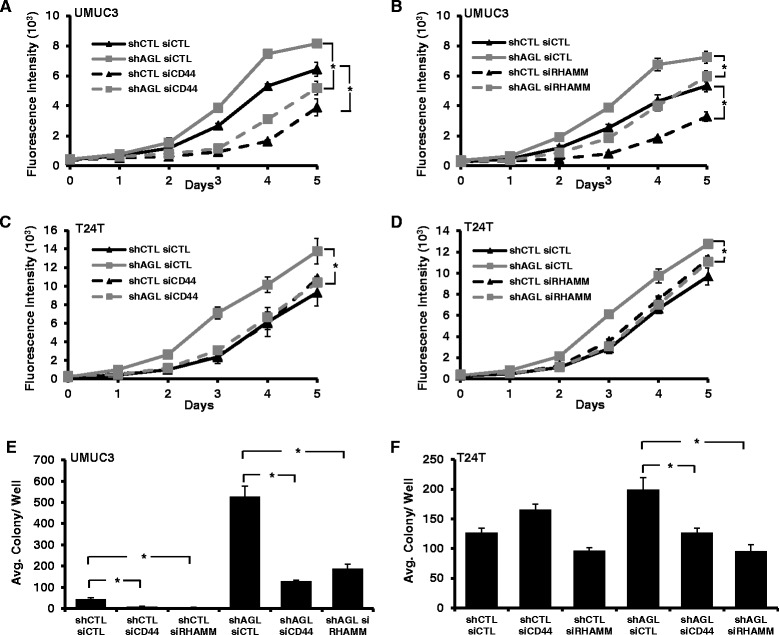
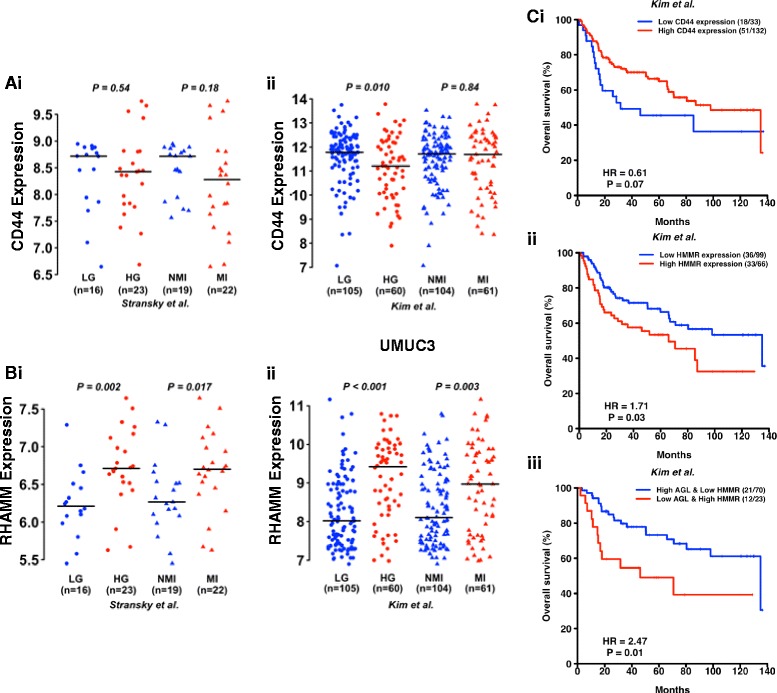
Results: Higher amounts of cleaved Cas3, Cas9 and PARP was observed in AGL low bladder cancer cell with loss of HAS2, CD44 or RHAMM. TUNEL staining showed more apoptotic cells with loss of HAS2, CD44 or RHAMM in AGL low bladder cancer cells. This revealed that bladder cancer cells whose aggressive growth is mediated by loss of AGL are susceptible to apoptosis with loss of HAS2, CD44 or RHAMM. Interestingly loss of either CD44 or RHAMM induces apoptosis in different low AGL expressing bladder cancer cell lines. Growth assays showed that loss of CD44 and RHAMM predominantly inhibit anchorage dependent and independent growth of AGL low bladder cancer cells. Clinicopathologic analysis revealed that high RHAMM mRNA expression is a marker of poor patient outcome in bladder cancer and patients with high RHAMM and low AGL tumor mRNA expression have poor survival.
Conclusion: Our findings strongly point to the importance of the HAS2-HA-CD44/RHAMM pathway for rapid growth of bladder cancer cells with loss of AGL and provides rational for targeting this pathway at various steps for "personalized" treatment of bladder cancer patients based of their AGL expression status.
Keywords: AGL; Bladder cancer; CD44; HAS2; RHAMM.
Figures
References
-
- Murray RK, Granner DK, Mayes PA, Rodwell VW. Harper's Illustrated Biochemistry. 26. New York: McGraw Hill; 2003. p. 180.
-
- Kishnani PS, Austin SL, Arn P, Bali DS, Boney A, Case LE, Chung WK, Desai DM, El-Gharbawy A, Haller R, Smit GP, Smith AD, Hobson-Webb LD, Wechsler SB, Weinstein DA, Watson MS. Glycogen storage disease type III diagnosis and management guidelines. Genet Med. 2010;12(7):446–63. doi: 10.1097/GIM.0b013e3181e655b6. - DOI - PubMed
-
- Guin S, Pollard C, Ru Y, Ritterson Lew C, Duex JE, Dancik G, Owens C, Spencer A, Knight S, Holemon H, Gupta S, Hansel D, Hellerstein M, Lorkiewicz P, Lane AN, Fan TW, Theodorescu D. Role in tumor growth of a glycogen debranching enzyme lost in glycogen storage disease. J Natl Cancer Inst. 2014;106:5. doi: 10.1093/jnci/dju062. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
