

Denys-Drash syndrome associated WT1 glutamine 369 mutants have altered sequence-preferences and altered responses to epigenetic modifications
- PMID: 27596598
- PMCID: PMC5137435
- DOI: 10.1093/nar/gkw766
Denys-Drash syndrome associated WT1 glutamine 369 mutants have altered sequence-preferences and altered responses to epigenetic modifications
Abstract
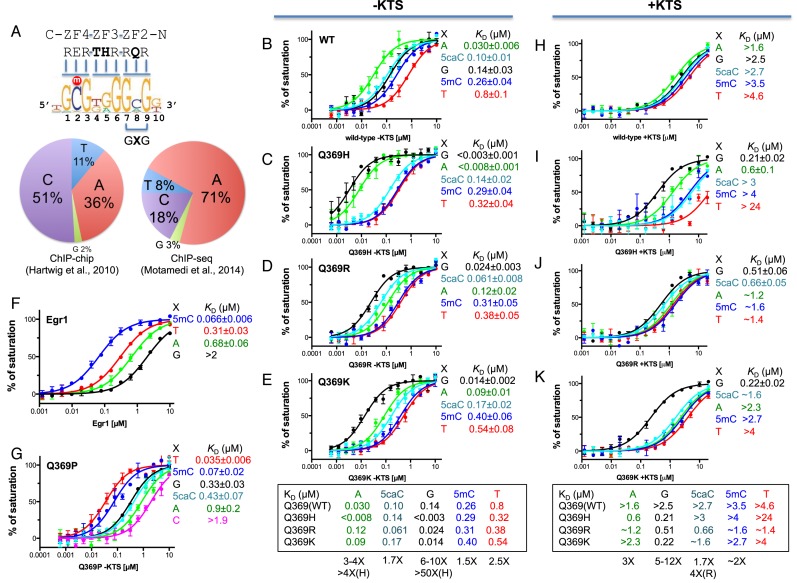
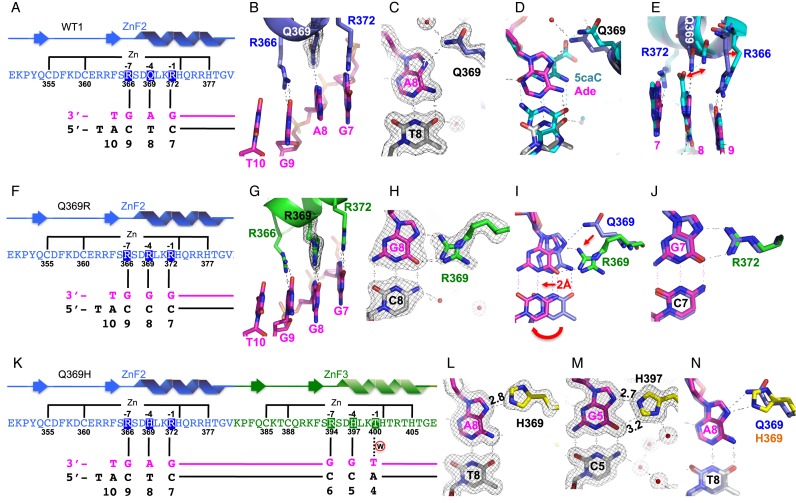
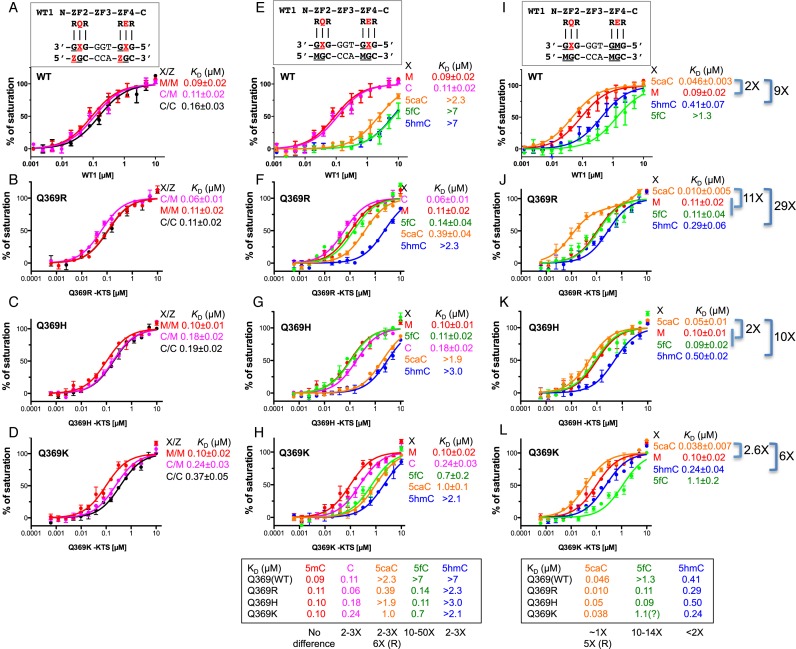
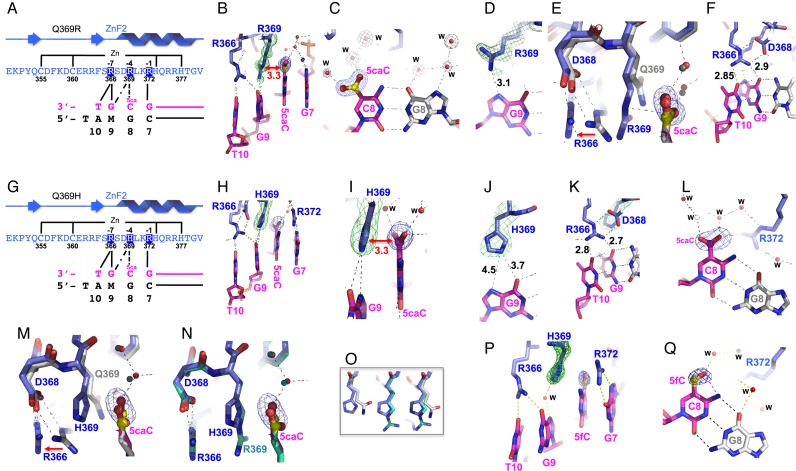
Mutations in human zinc-finger transcription factor WT1 result in abnormal development of the kidneys and genitalia and an array of pediatric problems including nephropathy, blastoma, gonadal dysgenesis and genital discordance. Several overlapping phenotypes are associated with WT1 mutations, including Wilms tumors, Denys-Drash syndrome (DDS), Frasier syndrome (FS) and WAGR syndrome (Wilms tumor, aniridia, genitourinary malformations, and mental retardation). These conditions vary in severity from individual to individual; they can be fatal in early childhood, or relatively benign into adulthood. DDS mutations cluster predominantly in zinc fingers (ZF) 2 and 3 at the C-terminus of WT1, which together with ZF4 determine the sequence-specificity of DNA binding. We examined three DDS associated mutations in ZF2 of human WT1 where the normal glutamine at position 369 is replaced by arginine (Q369R), lysine (Q369K) or histidine (Q369H). These mutations alter the sequence-specificity of ZF2, we find, changing its affinity for certain bases and certain epigenetic forms of cytosine. X-ray crystallography of the DNA binding domains of normal WT1, Q369R and Q369H in complex with preferred sequences revealed the molecular interactions responsible for these affinity changes. DDS is inherited in an autosomal dominant fashion, implying a gain of function by mutant WT1 proteins. This gain, we speculate, might derive from the ability of the mutant proteins to sequester WT1 into unproductive oligomers, or to erroneously bind to variant target sequences.
© The Author(s) 2016. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
Role for first zinc finger of WT1 in DNA sequence specificity: Denys-Drash syndrome-associated WT1 mutant in ZF1 enhances affinity for a subset of WT1 binding sites.Nucleic Acids Res. 2018 May 4;46(8):3864-3877. doi: 10.1093/nar/gkx1274. Nucleic Acids Res. 2018. PMID: 29294058 Free PMC article.
-
An unusual phenotype of Frasier syndrome due to IVS9 +4C>T mutation in the WT1 gene: predominantly male ambiguous genitalia and absence of gonadal dysgenesis.J Clin Endocrinol Metab. 2002 Jun;87(6):2500-5. doi: 10.1210/jcem.87.6.8521. J Clin Endocrinol Metab. 2002. PMID: 12050205 Review.
-
Effects of Denys-Drash syndrome point mutations on the DNA binding activity of the Wilms' tumor suppressor protein WT1.Biochemistry. 1996 Sep 17;35(37):12070-6. doi: 10.1021/bi960758o. Biochemistry. 1996. PMID: 8810912
-
A review of the phenotypic variation due to the Denys-Drash syndrome-associated germline WT1 mutation R362X.Hum Mutat. 2002 Apr;19(4):462. doi: 10.1002/humu.9031. Hum Mutat. 2002. PMID: 11933209 Review.
-
A review of the genetic background in complicated WT1-related disorders.Clin Exp Nephrol. 2025 Jan;29(1):1-9. doi: 10.1007/s10157-024-02539-x. Epub 2024 Jul 13. Clin Exp Nephrol. 2025. PMID: 39002031 Free PMC article. Review.
Cited by
-
Suspicion of Frasier's Syndrome in the Nephrology Unit of the State University Hospital of Haiti: Case Study and Review of Literature.Int Med Case Rep J. 2021 Aug 12;14:533-538. doi: 10.2147/IMCRJ.S325619. eCollection 2021. Int Med Case Rep J. 2021. PMID: 34408503 Free PMC article.
-
Role for first zinc finger of WT1 in DNA sequence specificity: Denys-Drash syndrome-associated WT1 mutant in ZF1 enhances affinity for a subset of WT1 binding sites.Nucleic Acids Res. 2018 May 4;46(8):3864-3877. doi: 10.1093/nar/gkx1274. Nucleic Acids Res. 2018. PMID: 29294058 Free PMC article.
-
Hotspots mutational analysis of Wilms tumor 1 gene in acute myeloid leukaemia; prevalence and clinical correlation in North Indian population.Am J Blood Res. 2020 Oct 15;10(5):179-189. eCollection 2020. Am J Blood Res. 2020. PMID: 33224562 Free PMC article.
-
Wilms' Tumor Protein 1 and Enzymatic Oxidation of 5-Methylcytosine in Brain Tumors: Potential Perspectives.Front Cell Dev Biol. 2018 Mar 22;6:26. doi: 10.3389/fcell.2018.00026. eCollection 2018. Front Cell Dev Biol. 2018. PMID: 29623275 Free PMC article. Review.
-
Molecular update on biology of Wilms Tumor 1 gene and its applications in acute myeloid leukemia.Am J Blood Res. 2020 Oct 15;10(5):151-160. eCollection 2020. Am J Blood Res. 2020. PMID: 33224559 Free PMC article. Review.
References
-
- Lee S.B., Haber D.A. Wilms tumor and the WT1 gene. Exp. Cell Res. 2001;264:74–99. - PubMed
-
- Wagner K.D., Wagner N., Schedl A. The complex life of WT1. J. Cell Sci. 2003;116:1653–1658. - PubMed
-
- Niaudet P., Gubler M.C. WT1 and glomerular diseases. Pediatric Nephrol. 2006;21:1653–1660. - PubMed
-
- Wang Z.Y., Qiu Q.Q., Deuel T.F. The Wilms' tumor gene product WT1 activates or suppresses transcription through separate functional domains. J. Biol. Chem. 1993;268:9172–9175. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
