

Horizontal Transfer of the Salmonella enterica Serovar Infantis Resistance and Virulence Plasmid pESI to the Gut Microbiota of Warm-Blooded Hosts
- PMID: 27601577
- PMCID: PMC5013300
- DOI: 10.1128/mBio.01395-16
Horizontal Transfer of the Salmonella enterica Serovar Infantis Resistance and Virulence Plasmid pESI to the Gut Microbiota of Warm-Blooded Hosts
Abstract
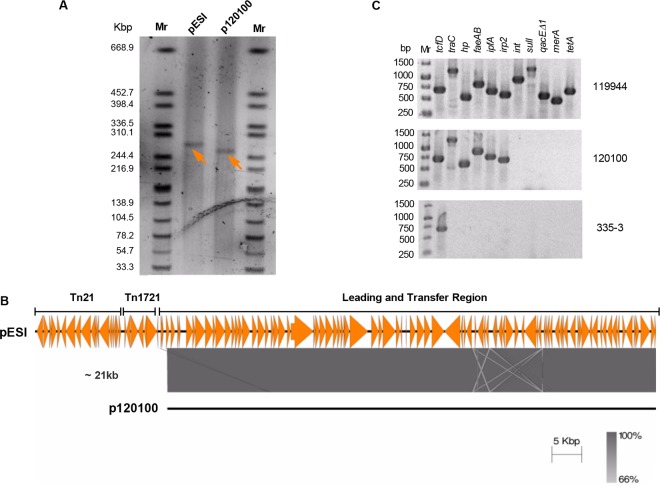
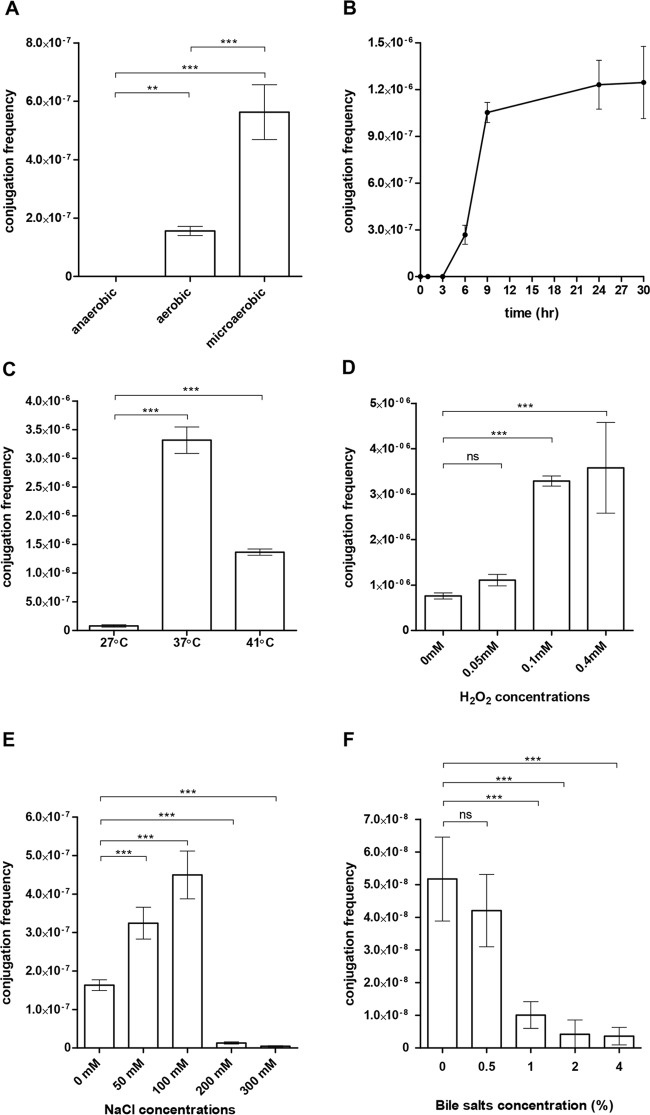
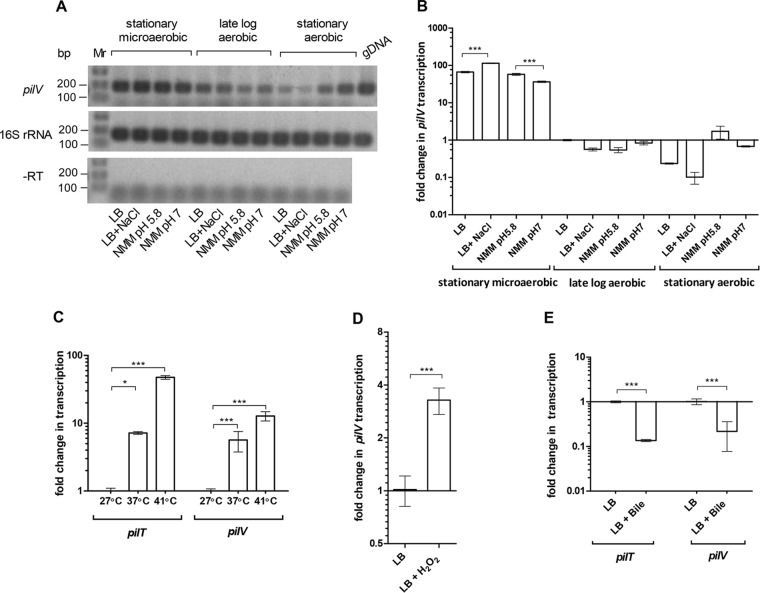
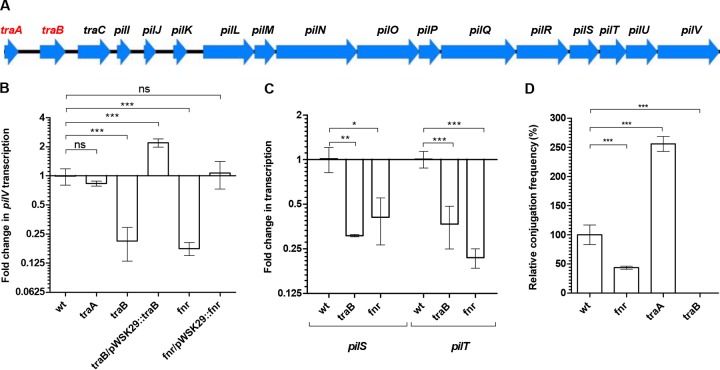
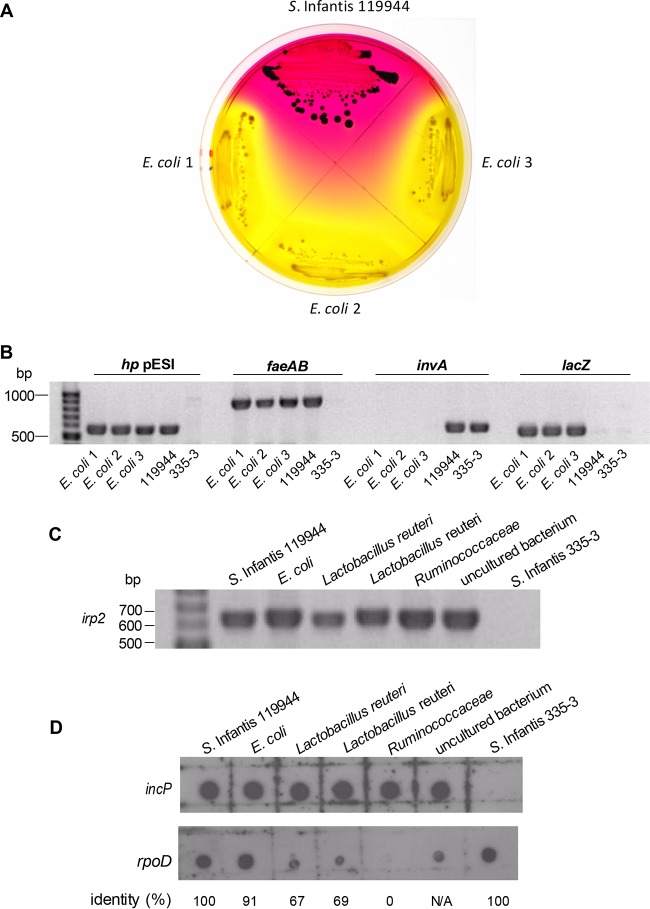
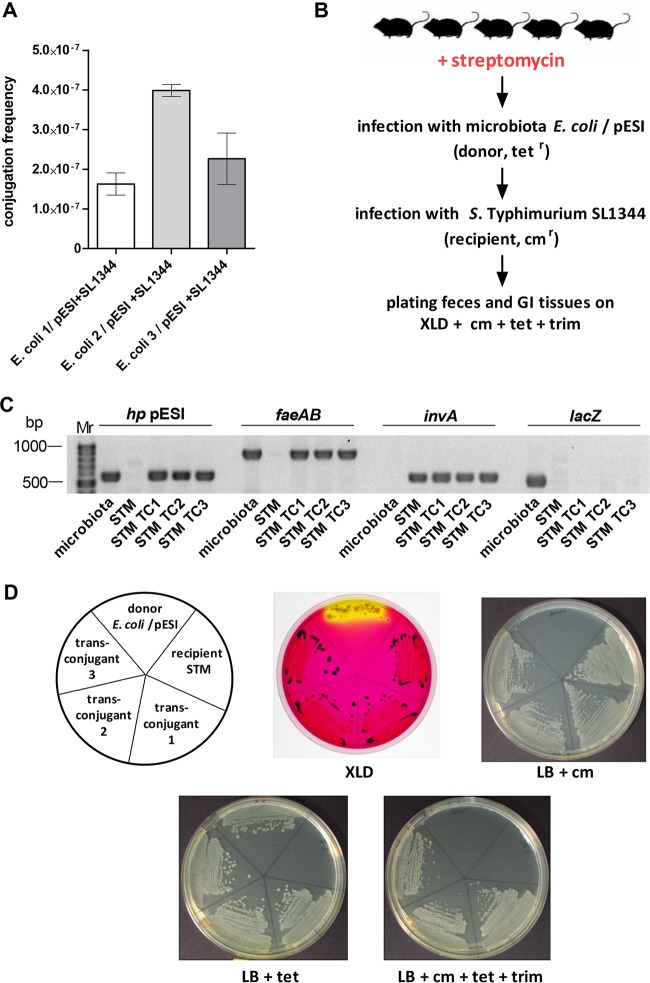
Salmonella enterica serovar Infantis is one of the prevalent salmonellae worldwide. Recently, we showed that the emergence of S Infantis in Israel was facilitated by the acquisition of a unique megaplasmid (pESI) conferring multidrug resistance and increased virulence phenotypes. Here we elucidate the ecology, transmission properties, and regulation of pESI. We show that despite its large size (~280 kb), pESI does not impose a significant metabolic burden in vitro and that it has been recently fixed in the domestic S Infantis population. pESI conjugation and the transcription of its pilus (pil) genes are inhibited at the ambient temperature (27°C) and by ≥1% bile but increased under temperatures of 37 to 41°C, oxidative stress, moderate osmolarity, and the microaerobic conditions characterizing the intestinal environment of warm-blooded animals. The pESI-encoded protein TraB and the oxygen homeostasis regulator Fnr were identified as transcriptional regulators of pESI conjugation. Using the mouse model, we show that following S Infantis infection, pESI can be horizontally transferred to the gut microbiota, including to commensal Escherichia coli strains. Possible transfer, but not persistence, of pESI was also observed into Gram-positive mouse microbiota species, especially Lactobacillus reuteri Moreover, pESI was demonstrated to further disseminate from gut microbiota to S. enterica serovar Typhimurium, in the context of gastrointestinal infection. These findings exhibit the ability of a selfish clinically relevant megaplasmid to distribute to and from the microbiota and suggest an overlooked role of the microbiota as a reservoir of mobile genetic elements and intermediator in the spread of resistance and virulence genes between commensals and pathogenic bacteria.
Importance: Plasmid conjugation plays a key role in microbial evolution, enabling the acquisition of new phenotypes, including resistance and virulence. Salmonella enterica serovar Infantis is one of the ubiquitous salmonellae worldwide and a major cause of foodborne infections. Previously, we showed that the emergence of S Infantis in Israel has involved the acquisition of a unique megaplasmid (pESI) conferring multidrug resistance and increased virulence phenotypes. Recently, the emergence of another S Infantis strain carrying a pESI-like plasmid was identified in Italy, suggesting that the acquisition of pESI may be common to different emergent S Infantis populations globally. Transmission of this plasmid to other strains or bacterial species is an alarming scenario. Understanding the ecology, regulation, and transmission properties of clinically relevant plasmids and the role of the microbiota in their spreading offers a new mechanism explaining the emergence of new pathogenic and resistant biotypes and may assist in the development of appropriate surveillance and prevention measures.
Copyright © 2016 Aviv et al.
Figures
References
-
- Galanis E, Lo Fo Wong DM, Patrick ME, Binsztein N, Cieslik A, Chalermchikit T, Aidara-Kane A, Ellis A, Angulo FJ, Wegener HC, World Health Organization Global Salm-Surv . 2006. Web-based surveillance and global Salmonella distribution, 2000–2002. Emerg Infect Dis 12:381–388. doi: 10.3201/eid1205.050854. - DOI - PMC - PubMed
-
- Majowicz SE, Musto J, Scallan E, Angulo FJ, Kirk M, O’Brien SJ, Jones TF, Fazil A, Hoekstra RM, International Collaboration on Enteric Disease Burden of Illness Studies . 2010. The global burden of nontyphoidal Salmonella gastroenteritis. Clin Infect Dis 50:882–889. doi: 10.1086/650733. - DOI - PubMed
-
- Crim SM, Griffin PM, Tauxe R, Marder EP, Gilliss D, Conquist AB, Cartter M, Tobin-D’Angelo M, Blythe D, Smith K, Lathrop S, Zansky S, Cieslak PR, Dunn J, Holt KG, Wolpert B, Henao OL, Centers for Disease Control and Prevention . 2015. Preliminary incidence and trends of infection with pathogens transmitted commonly through food—Foodborne Diseases Active Surveillance Network, 10 U.S. sites, 2006–2014. MMWR Morb Mortal Wkly Rep 64:495–499. - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
