

In B cells, phosphatidylinositol 5-phosphate 4-kinase-α synthesizes PI(4,5)P2 to impact mTORC2 and Akt signaling
- PMID: 27601656
- PMCID: PMC5035885
- DOI: 10.1073/pnas.1522478113
In B cells, phosphatidylinositol 5-phosphate 4-kinase-α synthesizes PI(4,5)P2 to impact mTORC2 and Akt signaling
Abstract
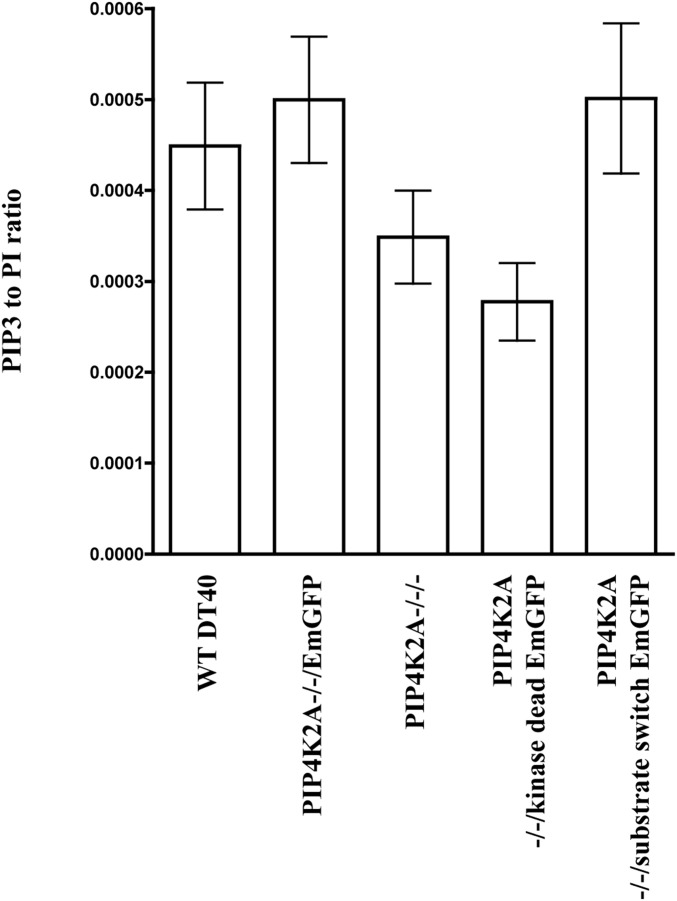
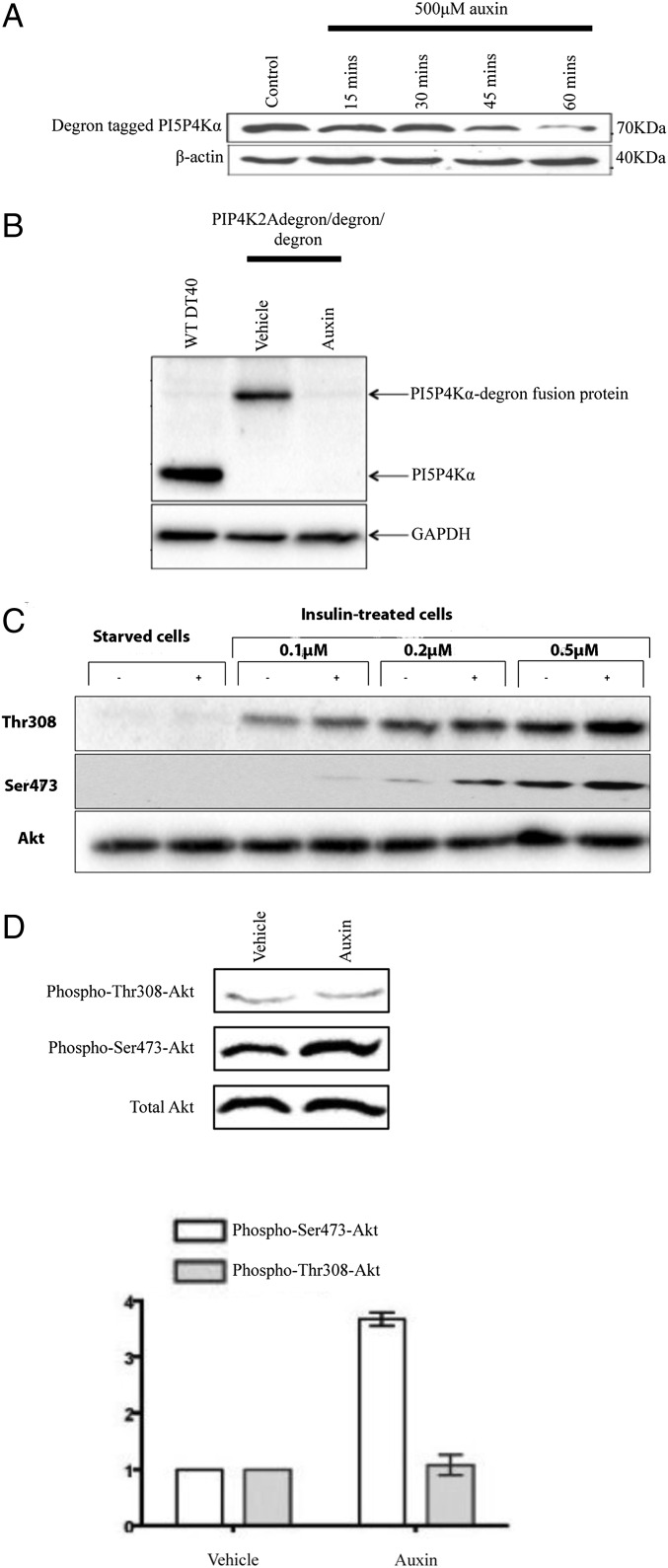
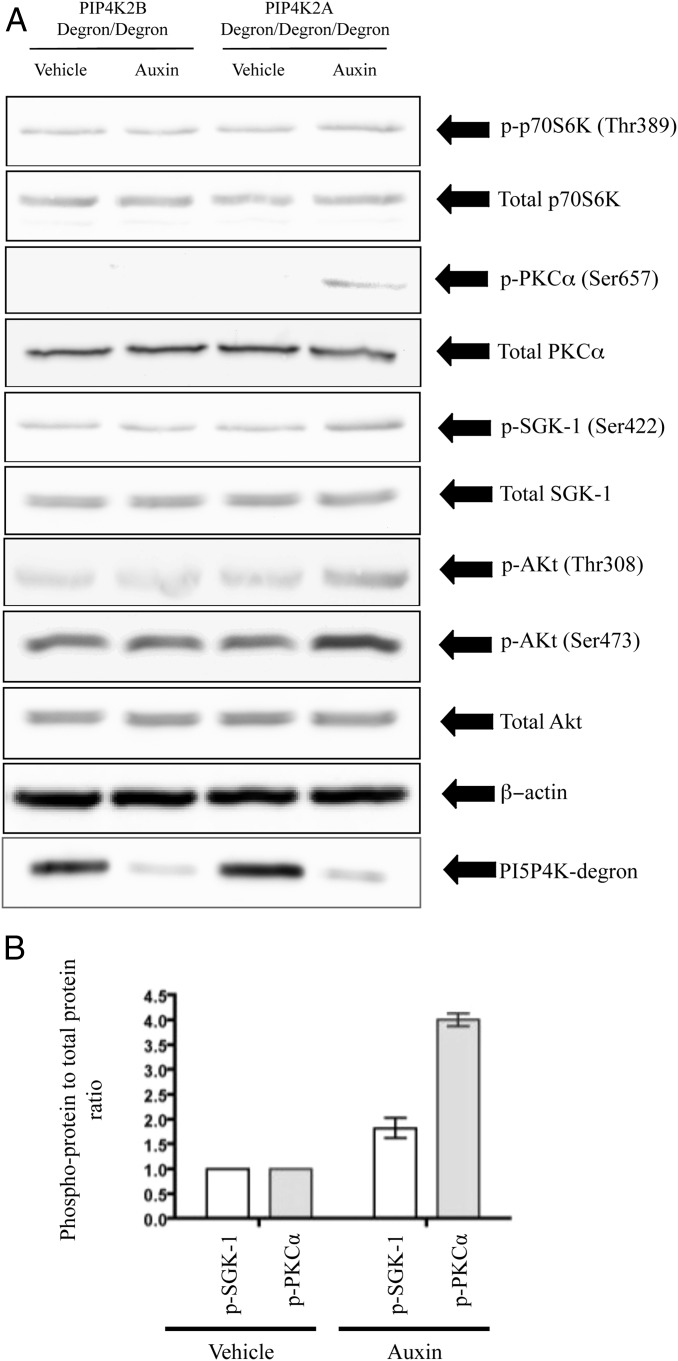
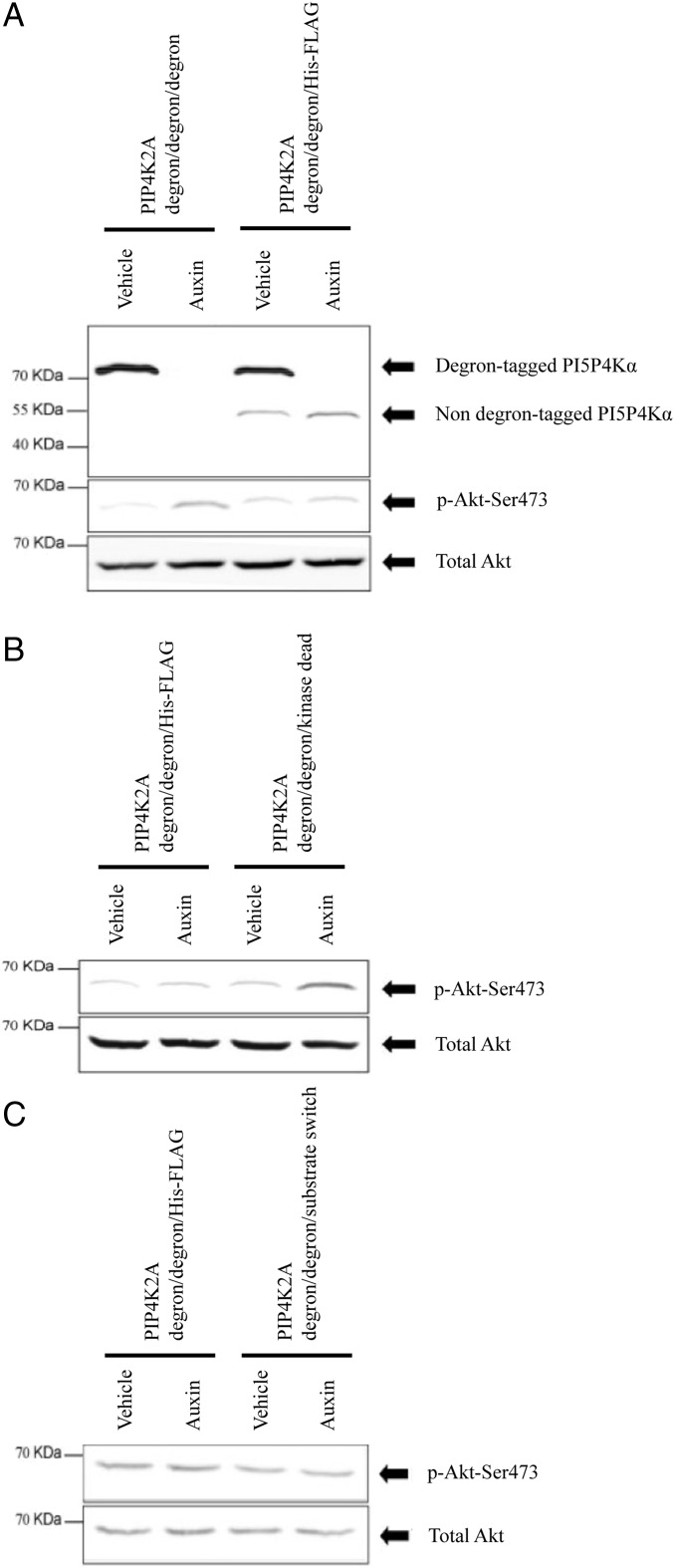
Phosphatidylinositol 5-phosphate 4-kinases (PI5P4Ks) are enigmatic lipid kinases with physiological functions that are incompletely understood, not the least because genetic deletion and cell transfection have led to contradictory data. Here, we used the genetic tractability of DT40 cells to create cell lines in which endogenous PI5P4Kα was removed, either stably by genetic deletion or transiently (within 1 h) by tagging the endogenous protein genomically with the auxin degron. In both cases, removal impacted Akt phosphorylation, and by leaving one PI5P4Kα allele present but mutating it to be kinase-dead or have PI4P 5-kinase activity, we show that all of the effects on Akt phosphorylation were dependent on the ability of PI5P4Kα to synthesize phosphatidylinositol (4,5)-bisphosphate [PI(4,5)P2] rather than to remove PI5P. Although stable removal of PI5P4Kα resulted in a pronounced decrease in Akt phosphorylation at Thr308 and Ser473, in part because of reduced plasma membrane PIP3, its acute removal led to an increase in Akt phosphorylation only at Ser473. This process invokes activation primarily of mammalian target of rapamycin complex 2 (mTORC2), which was confirmed by increased phosphorylation of other mTORC2 substrates. These findings establish PI5P4Kα as a kinase that synthesizes a physiologically relevant pool of PI(4,5)P2 and as a regulator of mTORC2, and show a phenomenon similar to the "butterfly effect" described for phosphatidylinositol 3-kinase Iα [Hart JR, et al. (2015) Proc Natl Acad Sci USA 112(4):1131-1136], whereby through apparently the same underlying mechanism, the removal of a protein's activity from a cell can have widely divergent effects depending on the time course of that removal.
Keywords: Akt; mTOR; phosphatidylinositol (4,5)-bisphosphate; phosphatidylinositol 5-phosphate; phosphatidylinositol 5-phosphate 4-kinase.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Viaud J, Boal F, Tronchère H, Gaits-Iacovoni F, Payrastre B. Phosphatidylinositol 5-phosphate: A nuclear stress lipid and a tuner of membranes and cytoskeleton dynamics. BioEssays. 2014;36(3):260–272. - PubMed
-
- Rozenvayn N, Flaumenhaft R. Phosphatidylinositol 4,5-bisphosphate mediates Ca2+-induced platelet alpha-granule secretion: Evidence for type II phosphatidylinositol 5-phosphate 4-kinase function. J Biol Chem. 2001;276(25):22410–22419. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
