

Diversity-oriented synthesis yields novel multistage antimalarial inhibitors
- PMID: 27602946
- PMCID: PMC5515376
- DOI: 10.1038/nature19804
Diversity-oriented synthesis yields novel multistage antimalarial inhibitors
Erratum in
-
Author Correction: Diversity-oriented synthesis yields novel multistage antimalarial inhibitors.Nature. 2025 Dec;648(8094):E12-E13. doi: 10.1038/s41586-025-09938-4. Nature. 2025. PMID: 41315741 No abstract available.
Abstract
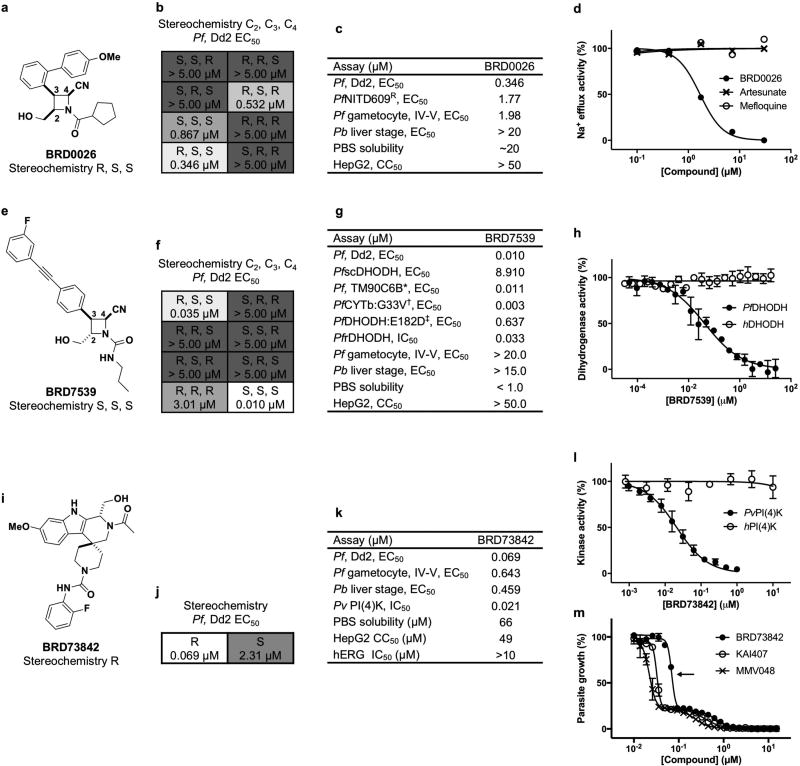
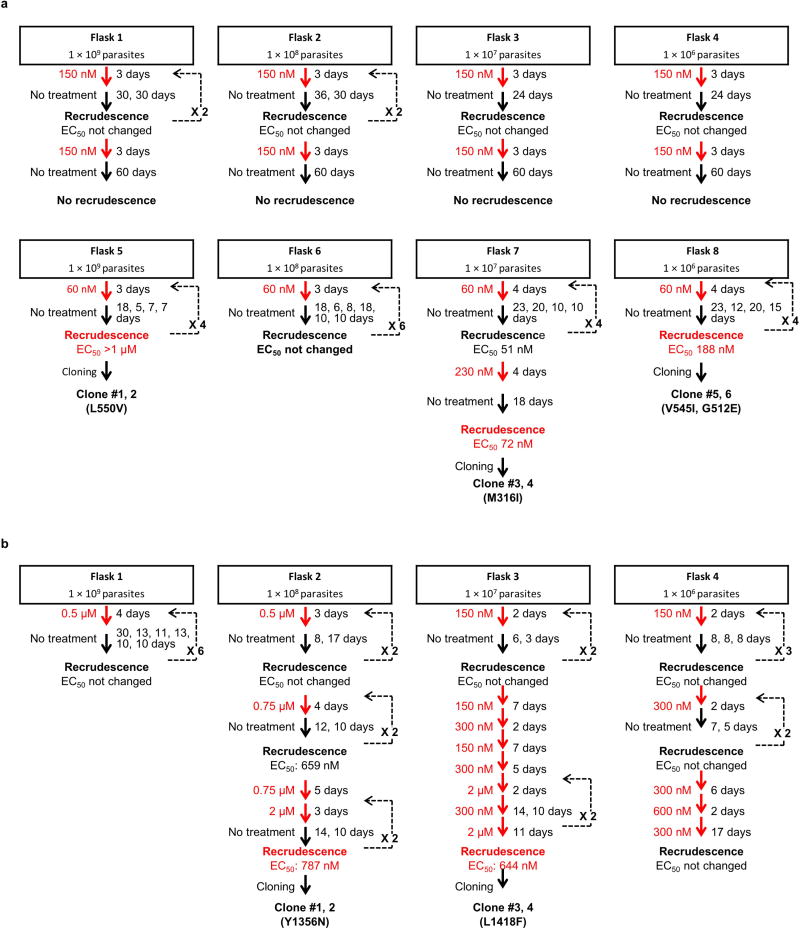
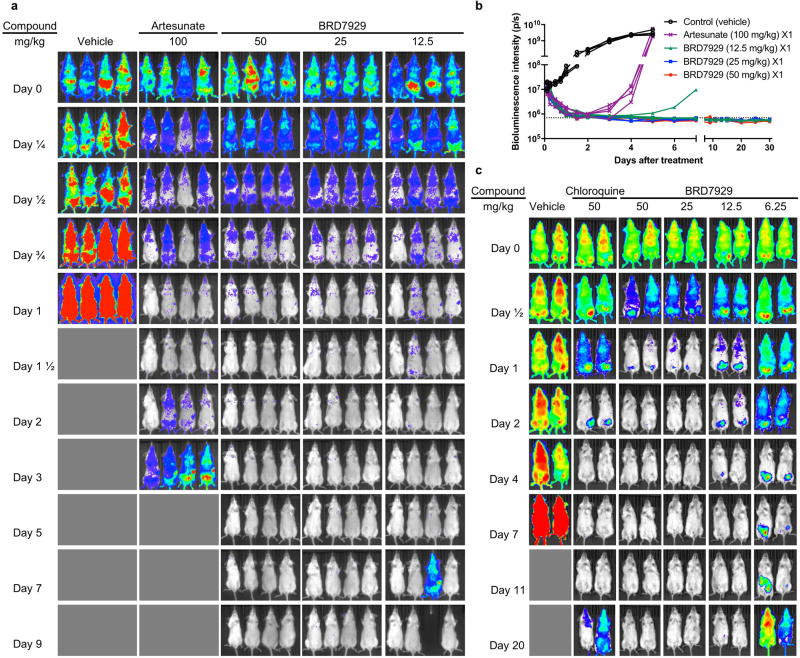
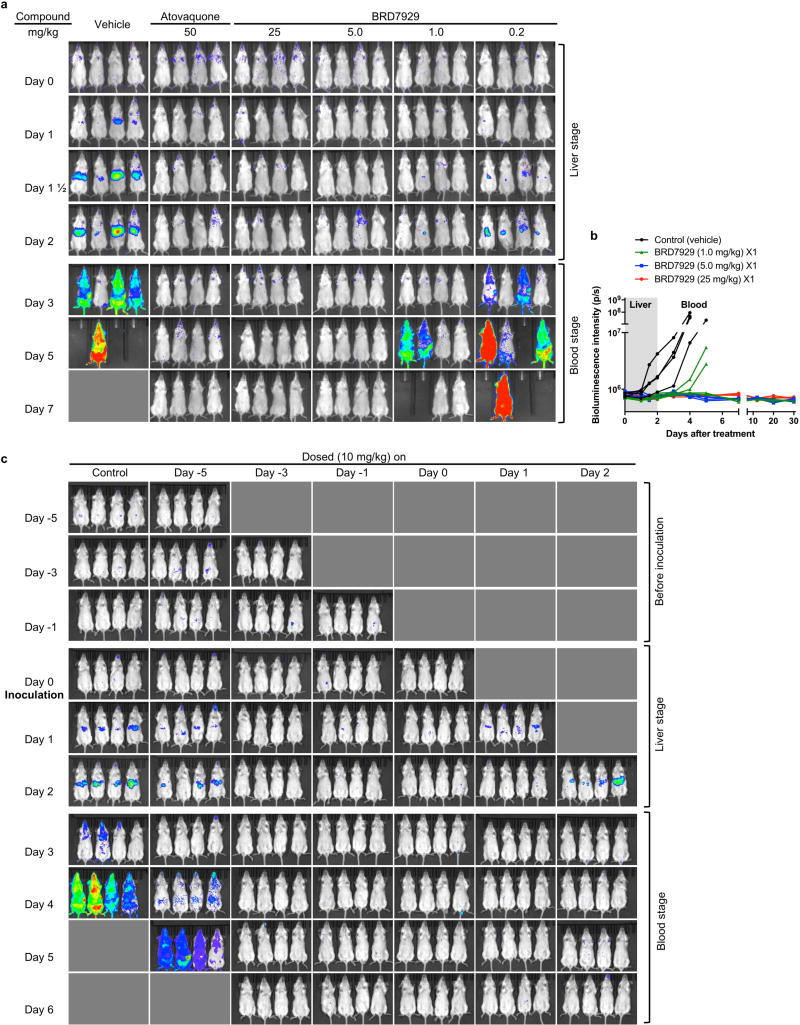
Antimalarial drugs have thus far been chiefly derived from two sources-natural products and synthetic drug-like compounds. Here we investigate whether antimalarial agents with novel mechanisms of action could be discovered using a diverse collection of synthetic compounds that have three-dimensional features reminiscent of natural products and are underrepresented in typical screening collections. We report the identification of such compounds with both previously reported and undescribed mechanisms of action, including a series of bicyclic azetidines that inhibit a new antimalarial target, phenylalanyl-tRNA synthetase. These molecules are curative in mice at a single, low dose and show activity against all parasite life stages in multiple in vivo efficacy models. Our findings identify bicyclic azetidines with the potential to both cure and prevent transmission of the disease as well as protect at-risk populations with a single oral dose, highlighting the strength of diversity-oriented synthesis in revealing promising therapeutic targets.
Figures
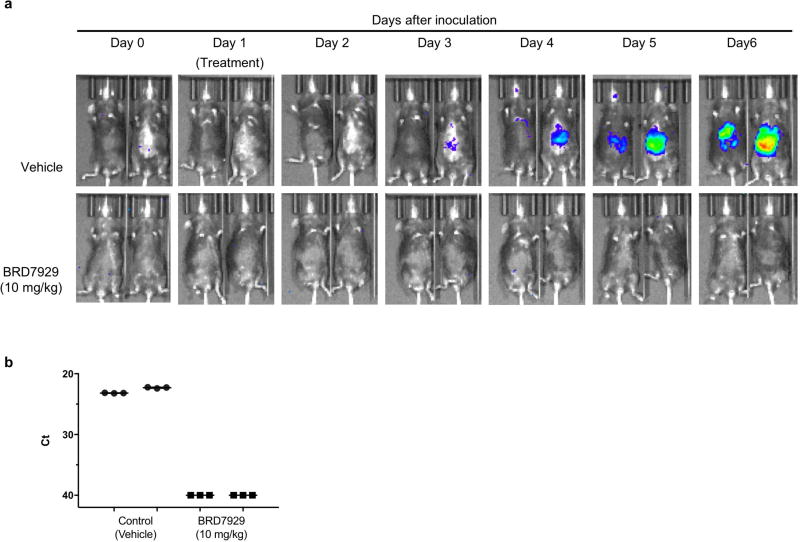
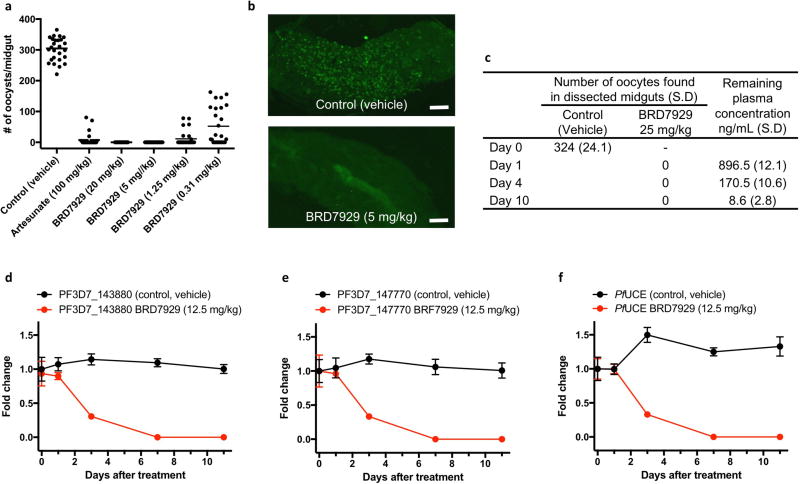
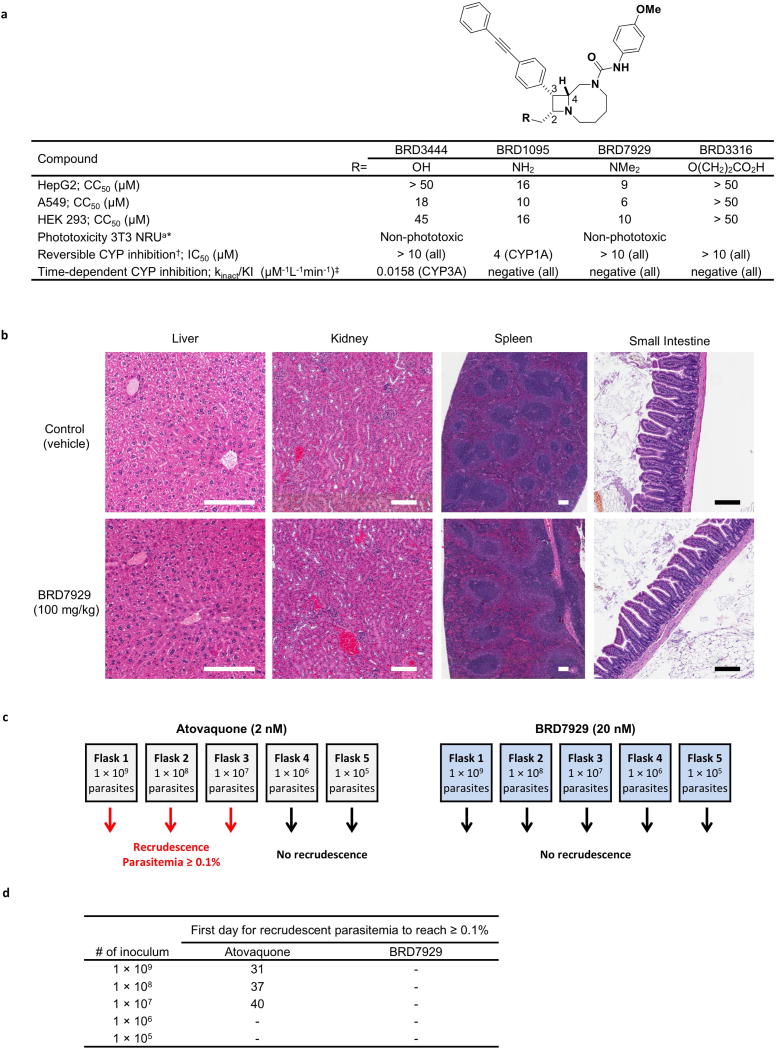
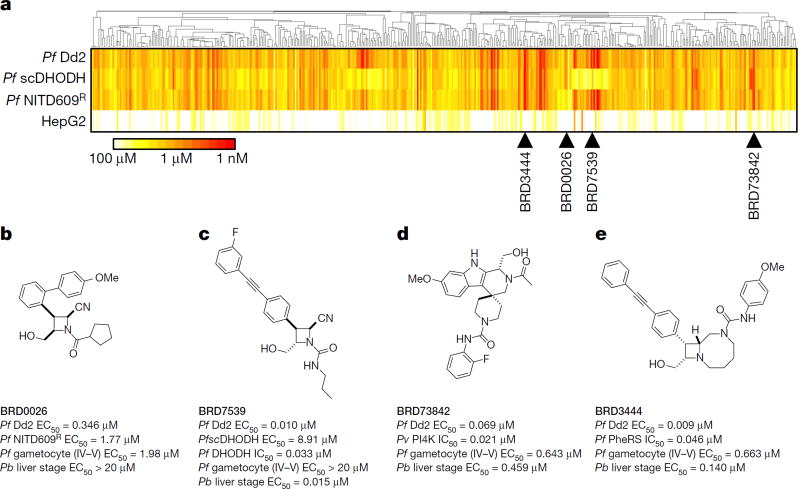
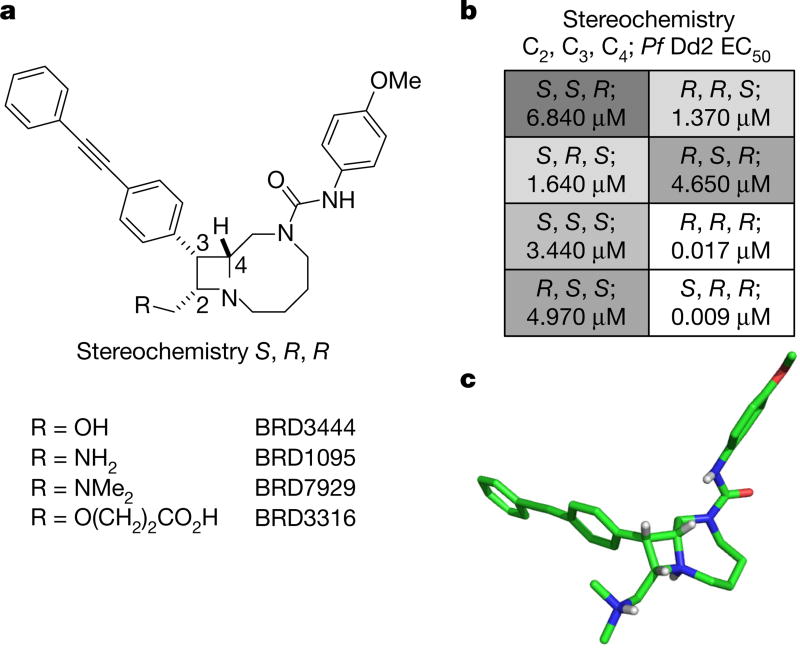
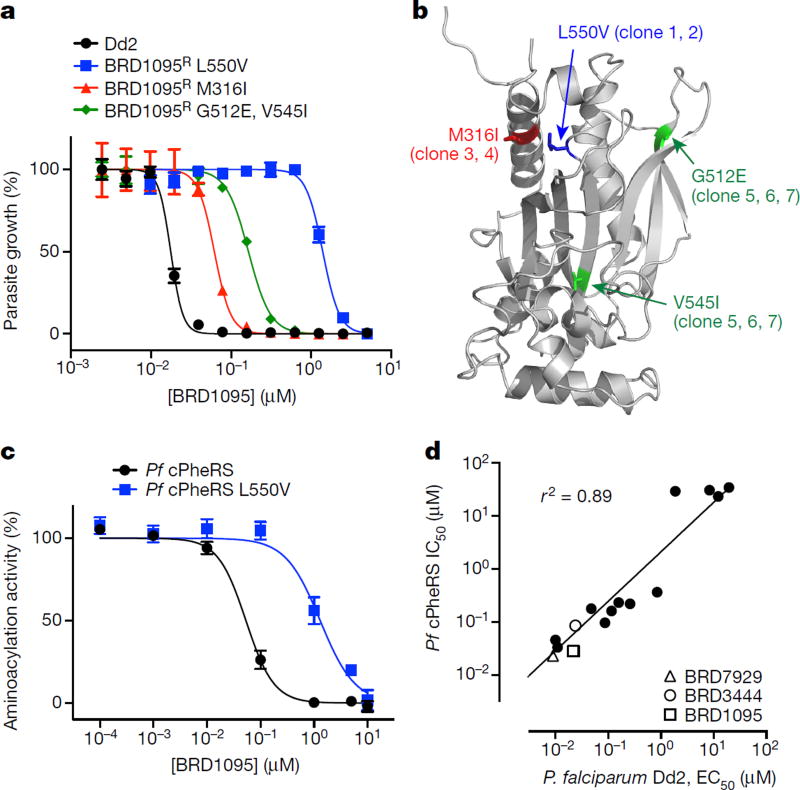
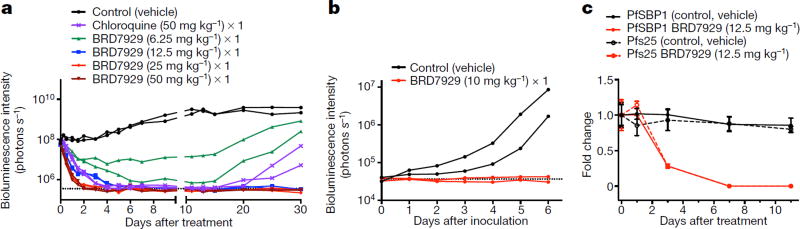
Comment in
-
Drug discovery: Chemical diversity targets malaria.Nature. 2016 Oct 20;538(7625):323-325. doi: 10.1038/nature19481. Epub 2016 Oct 12. Nature. 2016. PMID: 27732582 No abstract available.
-
Malaria: Novel antimalarial target identified.Nat Rev Drug Discov. 2016 Nov 3;15(11):750. doi: 10.1038/nrd.2016.218. Nat Rev Drug Discov. 2016. PMID: 27807362 No abstract available.
References
-
- Wells TN. Discovering and developing new medicines for malaria control and elimination. Infect. Disord. Drug Targets. 2013;13:292–302. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
