

Plasma membrane order and fluidity are diversely triggered by elicitors of plant defence
- PMID: 27604805
- PMCID: PMC5014163
- DOI: 10.1093/jxb/erw284
Plasma membrane order and fluidity are diversely triggered by elicitors of plant defence
Abstract
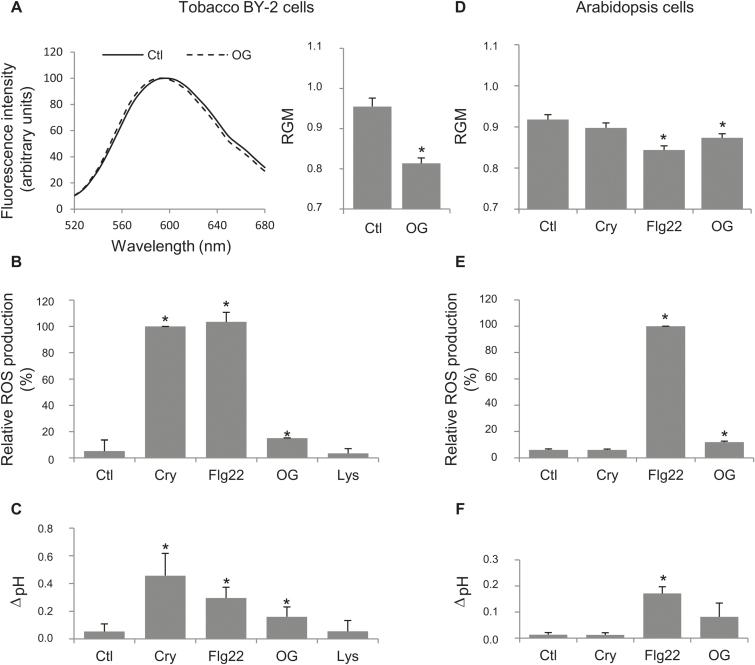
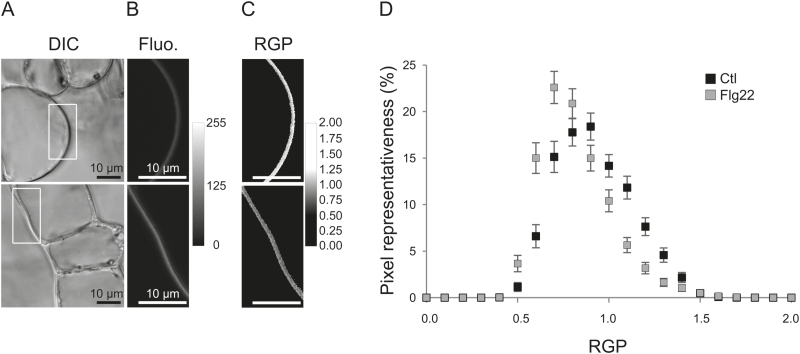
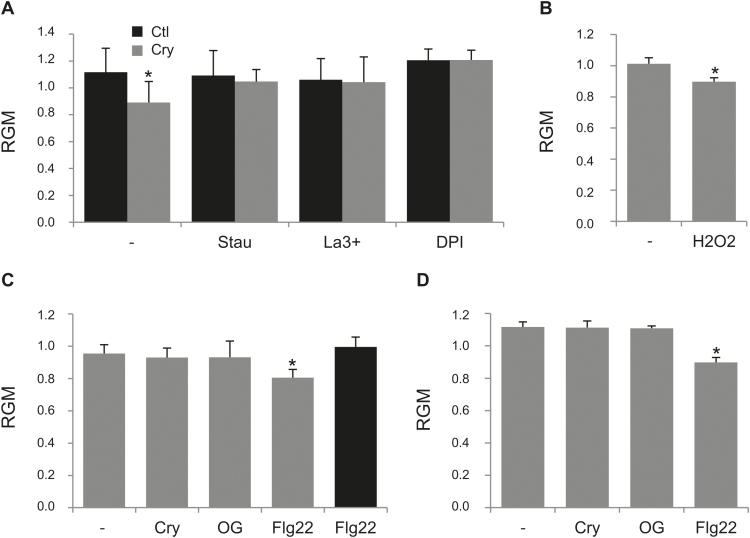
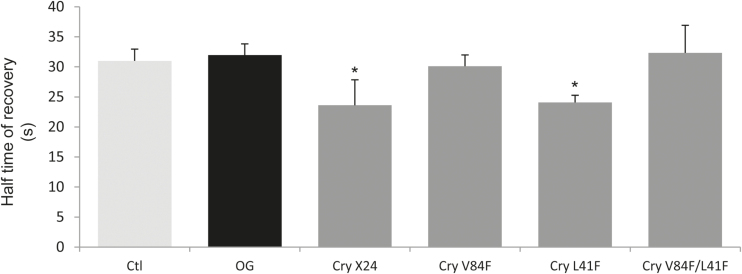
Although plants are exposed to a great number of pathogens, they usually defend themselves by triggering mechanisms able to limit disease development. Alongside signalling events common to most such incompatible interactions, modifications of plasma membrane (PM) physical properties could be new players in the cell transduction cascade. Different pairs of elicitors (cryptogein, oligogalacturonides, and flagellin) and plant cells (tobacco and Arabidopsis) were used to address the issue of possible modifications of plant PM biophysical properties induced by elicitors and their links to other events of the defence signalling cascade. We observed an increase of PM order whatever the elicitor/plant cell pair used, provided that a signalling cascade was induced. Such membrane modification is dependent on the NADPH oxidase-mediated reactive oxygen species production. Moreover, cryptogein, which is the sole elicitor able to trap sterols, is also the only one able to trigger an increase in PM fluidity. The use of cryptogein variants with altered sterol-binding properties confirms the strong correlation between sterol removal from the PM and PM fluidity enhancement. These results propose PM dynamics as a player in early signalling processes triggered by elicitors of plant defence.
Keywords: Cryptogein mutants; elicitors; fluidity; membrane order; plant defence; plasma membrane; reactive oxygen species; signalling..
© The Author 2016. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures
Similar articles
-
The plant defense elicitor cryptogein stimulates clathrin-mediated endocytosis correlated with reactive oxygen species production in bright yellow-2 tobacco cells.Plant Physiol. 2008 Mar;146(3):1255-66. doi: 10.1104/pp.107.111716. Epub 2008 Jan 9. Plant Physiol. 2008. PMID: 18184734 Free PMC article.
-
Long-chain bases and their phosphorylated derivatives differentially regulate cryptogein-induced production of reactive oxygen species in tobacco (Nicotiana tabacum) BY-2 cells.New Phytol. 2015 Feb;205(3):1239-1249. doi: 10.1111/nph.13094. Epub 2014 Oct 10. New Phytol. 2015. PMID: 25303640
-
Modification of plasma membrane organization in tobacco cells elicited by cryptogein.Plant Physiol. 2014 Jan;164(1):273-86. doi: 10.1104/pp.113.225755. Epub 2013 Nov 14. Plant Physiol. 2014. PMID: 24235133 Free PMC article.
-
Early signaling events induced by elicitors of plant defenses.Mol Plant Microbe Interact. 2006 Jul;19(7):711-24. doi: 10.1094/MPMI-19-0711. Mol Plant Microbe Interact. 2006. PMID: 16838784 Review.
-
Involvement of plasma membrane proteins in plant defense responses. Analysis of the cryptogein signal transduction in tobacco.Biochimie. 1999 Jun;81(6):663-8. doi: 10.1016/s0300-9084(99)80123-0. Biochimie. 1999. PMID: 10433120 Review.
Cited by
-
Synthetic Rhamnolipid Bolaforms trigger an innate immune response in Arabidopsis thaliana.Sci Rep. 2018 Jun 4;8(1):8534. doi: 10.1038/s41598-018-26838-y. Sci Rep. 2018. PMID: 29867089 Free PMC article.
-
Plant immune and growth receptors share common signalling components but localise to distinct plasma membrane nanodomains.Elife. 2017 Mar 6;6:e25114. doi: 10.7554/eLife.25114. Elife. 2017. PMID: 28262094 Free PMC article.
-
Membrane nanodomains and transport functions in plant.Plant Physiol. 2021 Dec 4;187(4):1839-1855. doi: 10.1093/plphys/kiab312. Plant Physiol. 2021. PMID: 35235669 Free PMC article. Review.
-
Interactions between lipids and proteins are critical for organization of plasma membrane-ordered domains in tobacco BY-2 cells.J Exp Bot. 2018 Jun 27;69(15):3545-3557. doi: 10.1093/jxb/ery152. J Exp Bot. 2018. PMID: 29722895 Free PMC article.
-
Plant defensive responses to insect eggs are inducible by general egg-associated elicitors.Sci Rep. 2024 Jan 11;14(1):1076. doi: 10.1038/s41598-024-51565-y. Sci Rep. 2024. PMID: 38212511 Free PMC article.
References
-
- Almagro L, Bru R, Pugin A, Pedreno MA. 2012. Early signaling network in tobacco cells elicited with methyl jasmonate and cyclodextrins. Plant Physiology and Biochemistry 51, 1–9. - PubMed
-
- Amari K, Diaz-Vivancos P, Pallas V, Sanchez-Pina MA, Hernandez JA. 2007. Oxidative stress induction by Prunus necrotic ringspot virus infection in apricot seeds. Physiologia Plantarum 131, 302–310. - PubMed
-
- Asai S, Yoshioka H. 2009. Nitric oxide as a partner of reactive oxygen species participates in disease resistance to nectrotophic pathogen Botryis cinerea in Nicotiana benthamiana. Molecular Plant-Microbe Interactions 22, 619–629. - PubMed
-
- Barna B, Fodor J, Harrach BD, Pogany M, Kiraly Z. 2012. The Janus face of reactive oxygen species in resistance and susceptibility of plants to necrotrophic and biotrophic pathogens. Plant Physiology and Biochemistry 59, 37–43. - PubMed
-
- Bartels S, Boller T. 2015. Quo vadis, Pep? Plant elicitor peptides at the crossroads of immunity, stress, and development. Journal of Experimental Botany 66, 5183–5193. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
