

The Genome as an Evolutionary Timepiece
- PMID: 27604881
- PMCID: PMC5630892
- DOI: 10.1093/gbe/evw220
The Genome as an Evolutionary Timepiece
Abstract
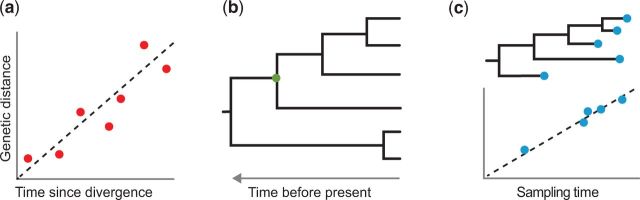
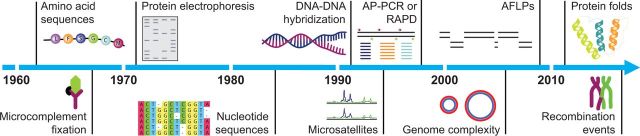
The molecular clock is a valuable and widely used tool for estimating evolutionary rates and timescales in biological research. There has been considerable progress in the theory and practice of molecular clocks over the past five decades. Although the idea of a molecular clock was originally put forward in the context of protein evolution and advanced using various biochemical techniques, it is now primarily applied to analyses of DNA sequences. An interesting but very underappreciated aspect of molecular clocks is that they can be based on genetic data other than DNA or protein sequences. For example, evolutionary timescales can be estimated using microsatellites, protein folds, and even the extent of recombination. These genome features hold great potential for molecular dating, particularly in cases where nucleotide sequences might be uninformative or unreliable. Here we present an outline of the different genetic data types that have been used for molecular dating, and we describe the features that good molecular clocks should possess. We hope that our article inspires further work on the genome as an evolutionary timepiece.
Keywords: evolutionary rate; genomic data; molecular clock; molecular dating; phylogenetic analysis.
© The Author 2016. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures
References
-
- Caetano-Anollés G, Wang M, Caetano-Anollés D, Mittenthal JE. 2009. The origin, evolution and structure of the protein world. Biochem J. 417:621–637. - PubMed
-
- Cohn VH, Thompson MA, Moore GP. 1984. Nucleotide sequence comparison of the Adh gene in three drosophilids. J Mol Evol. 20:31–37. - PubMed
-
- Davey JW, et al. 2011. Genome-wide genetic marker discovery and genotyping using next-generation sequencing. Nat Rev Genet. 12:499–510. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
