

Site-specific fluorescent labeling to visualize membrane translocation of a myristoyl switch protein
- PMID: 27605302
- PMCID: PMC5015116
- DOI: 10.1038/srep32866
Site-specific fluorescent labeling to visualize membrane translocation of a myristoyl switch protein
Abstract
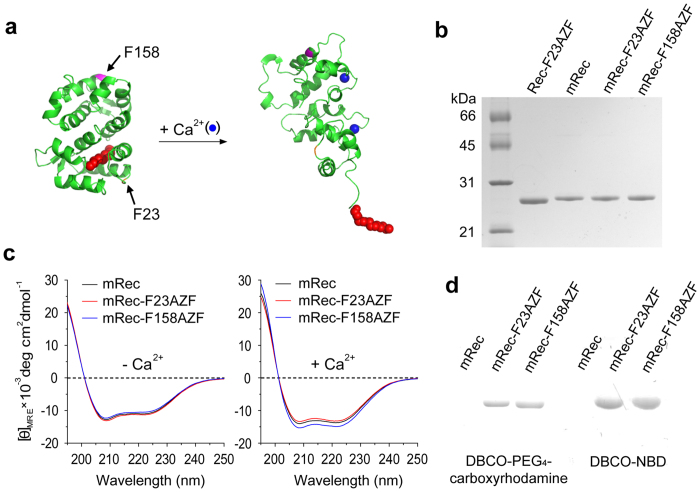
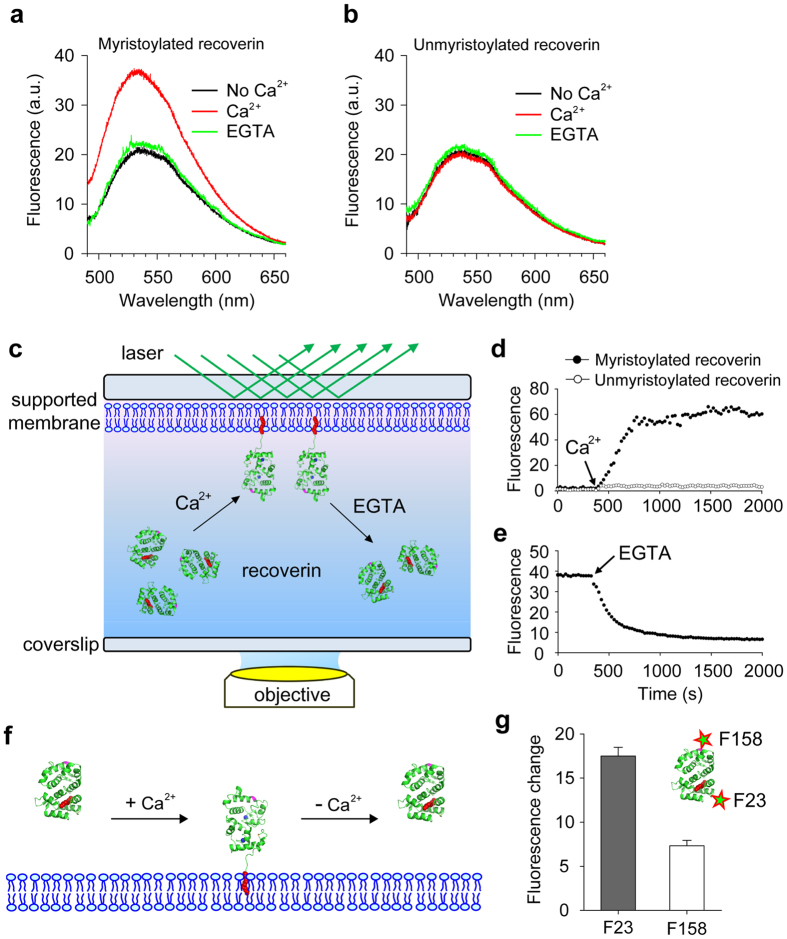
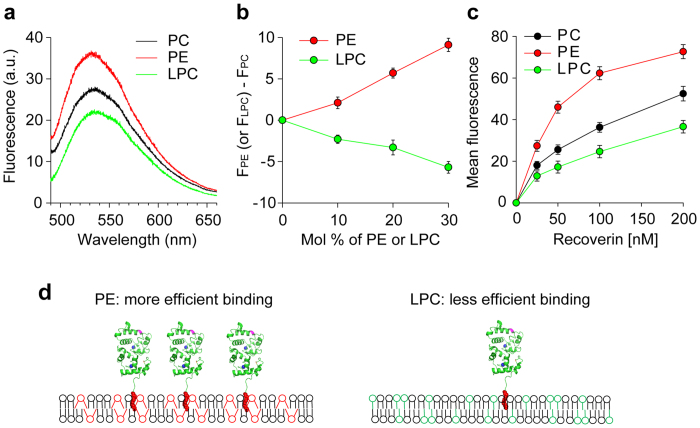
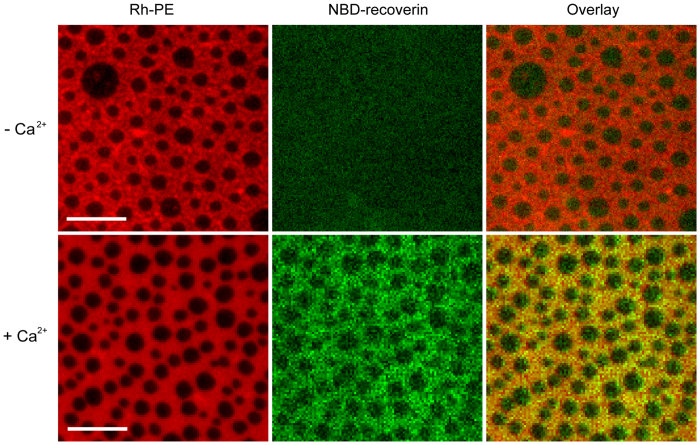
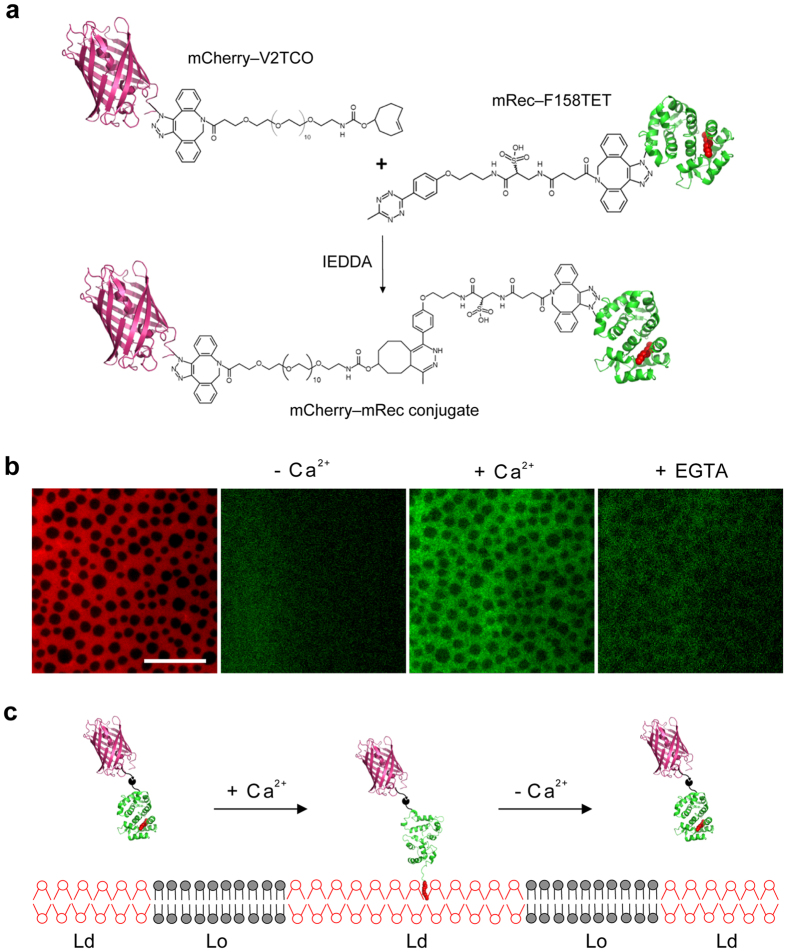
Fluorescence approaches have been widely used for elucidating the dynamics of protein-membrane interactions in cells and model systems. However, non-specific multi-site fluorescent labeling often results in a loss of native structure and function, and single cysteine labeling is not feasible when native cysteines are required to support a protein's folding or catalytic activity. Here, we develop a method using genetic incorporation of non-natural amino acids and bio-orthogonal chemistry to site-specifically label with a single fluorescent small molecule or protein the myristoyl-switch protein recoverin, which is involved in rhodopsin-mediated signaling in mammalian visual sensory neurons. We demonstrate reversible Ca(2+)-responsive translocation of labeled recoverin to membranes and show that recoverin favors membranes with negative curvature and high lipid fluidity in complex heterogeneous membranes, which confers spatio-temporal control over down-stream signaling events. The site-specific orthogonal labeling technique is promising for structural, dynamical, and functional studies of many lipid-anchored membrane protein switches.
Figures
References
-
- Weiss S. Fluorescence spectroscopy of single biomolecules. Science 283, 1676–1683 (1999). - PubMed
-
- Moerner W. E. & Fromm D. P. Methods of single-molecule fluorescence spectroscopy and microscopy. Rev Sci Instrum 74, 3597–3619 (2003).
-
- Joo C., Balci H., Ishitsuka Y., Buranachai C. & Ha T. Advances in single-molecule fluorescence methods for molecular biology. Annu Rev Biochem 77, 51–76 (2008). - PubMed
-
- Dizhoor A. M. et al. Recoverin: a calcium sensitive activator of retinal rod guanylate cyclase. Science 251, 915–918 (1991). - PubMed
-
- Gray-Keller M. P., Polans A. S., Palczewski K. & Detwiler P. B. The effect of recoverin-like calcium-binding proteins on the photoresponse of retinal rods. Neuron 10, 523–531 (1993). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
