

The Structural Architecture of an Infectious Mammalian Prion Using Electron Cryomicroscopy
- PMID: 27606840
- PMCID: PMC5015997
- DOI: 10.1371/journal.ppat.1005835
The Structural Architecture of an Infectious Mammalian Prion Using Electron Cryomicroscopy
Abstract
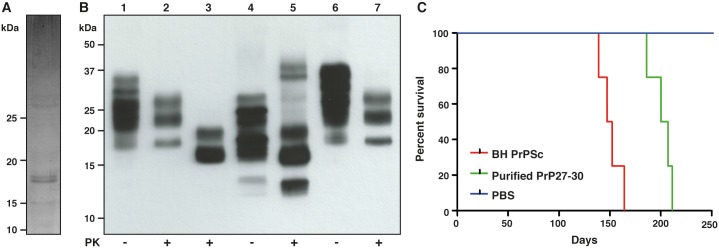
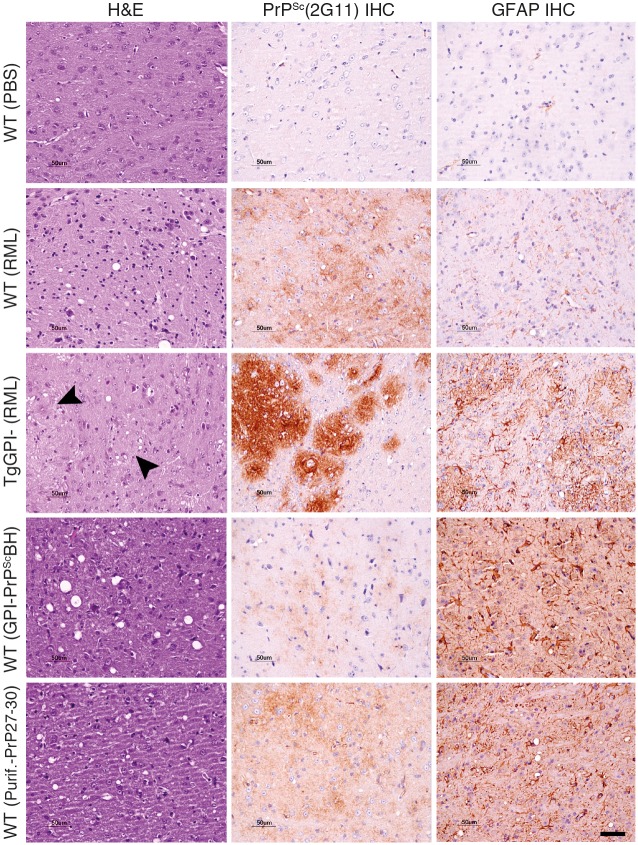
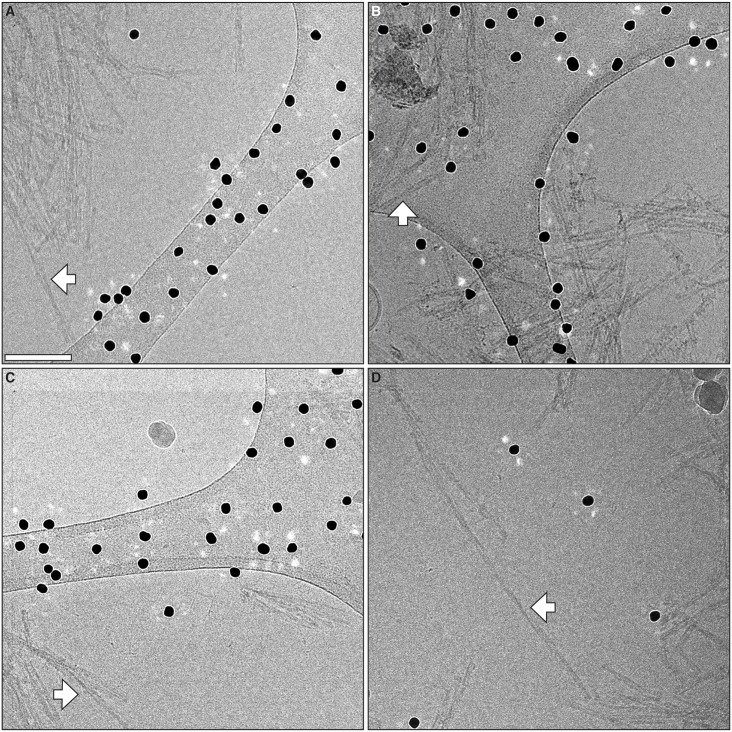
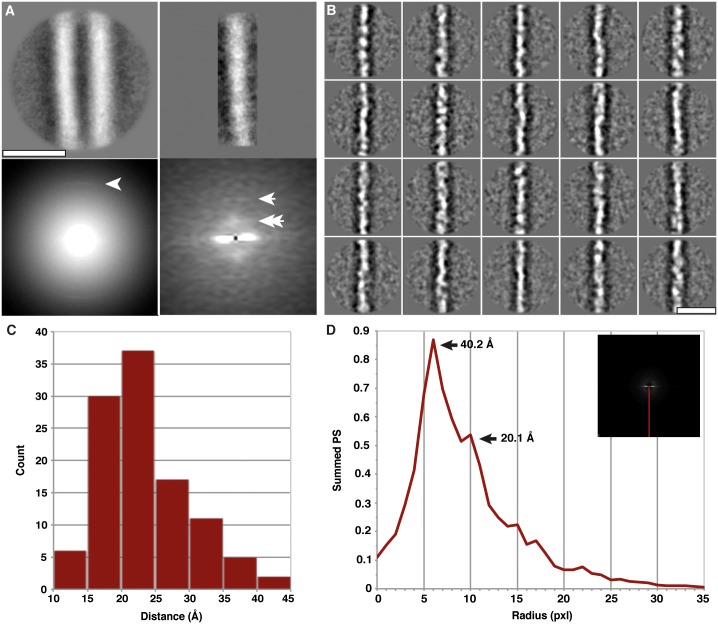
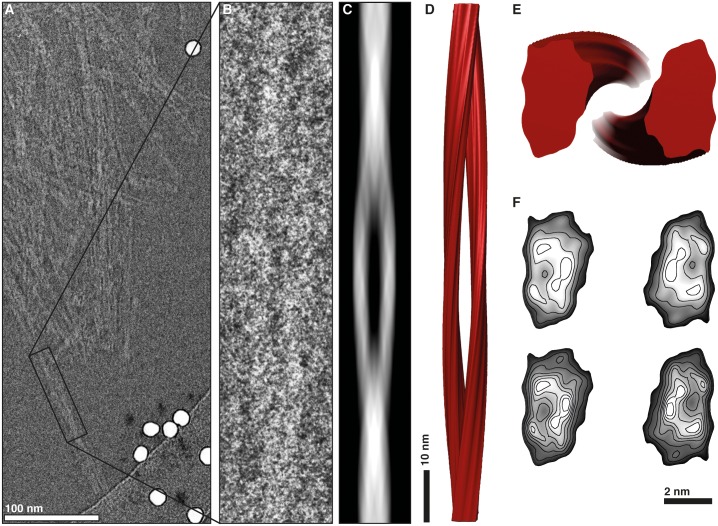
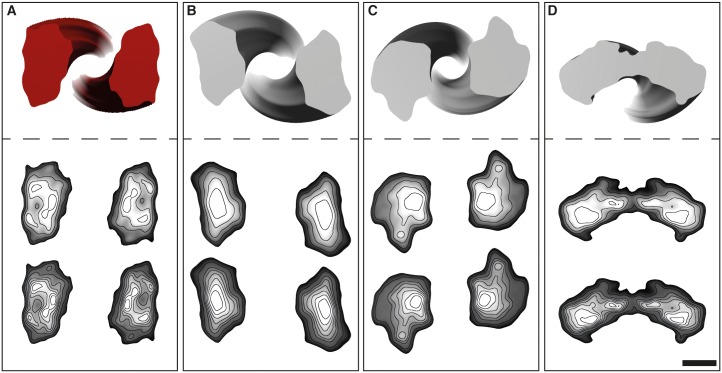
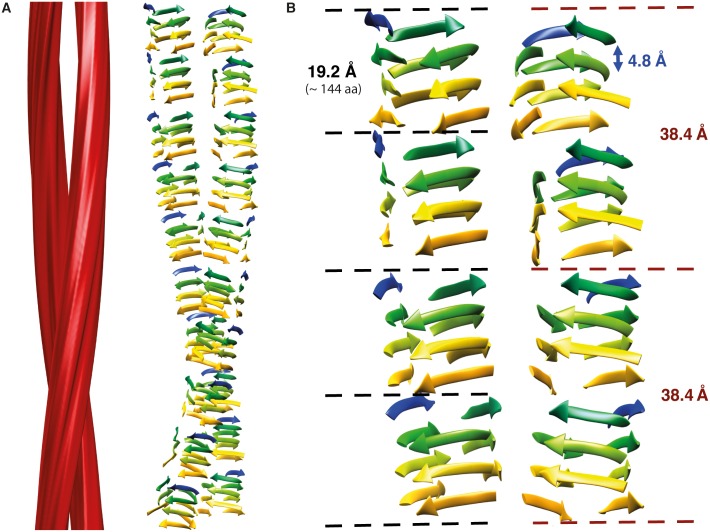
The structure of the infectious prion protein (PrPSc), which is responsible for Creutzfeldt-Jakob disease in humans and bovine spongiform encephalopathy, has escaped all attempts at elucidation due to its insolubility and propensity to aggregate. PrPSc replicates by converting the non-infectious, cellular prion protein (PrPC) into the misfolded, infectious conformer through an unknown mechanism. PrPSc and its N-terminally truncated variant, PrP 27-30, aggregate into amorphous aggregates, 2D crystals, and amyloid fibrils. The structure of these infectious conformers is essential to understanding prion replication and the development of structure-based therapeutic interventions. Here we used the repetitive organization inherent to GPI-anchorless PrP 27-30 amyloid fibrils to analyze their structure via electron cryomicroscopy. Fourier-transform analyses of averaged fibril segments indicate a repeating unit of 19.1 Å. 3D reconstructions of these fibrils revealed two distinct protofilaments, and, together with a molecular volume of 18,990 Å3, predicted the height of each PrP 27-30 molecule as ~17.7 Å. Together, the data indicate a four-rung β-solenoid structure as a key feature for the architecture of infectious mammalian prions. Furthermore, they allow to formulate a molecular mechanism for the replication of prions. Knowledge of the prion structure will provide important insights into the self-propagation mechanisms of protein misfolding.
Conflict of interest statement
MRV is an employee of FEI Company (Eindhoven, The Netherlands), this does not alter our adherence to all PLoS Pathogens policies on sharing data and materials.
Figures

References
-
- Riek R., Hornemann S., Wider G., Billeter M., Glockshuber R., and Wüthrich K. (1996). NMR structure of the mouse prion protein domain PrP(121–231). Nature 382, 180–182. - PubMed
-
- Knaus K.J., Morillas M., Swietnicki W., Malone M., Surewicz W.K., and Yee V.C. (2001). Crystal structure of the human prion protein reveals a mechanism for oligomerization. Nat. Struct. Biol. 8, 770–774. - PubMed
Publication types
MeSH terms
Substances
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
