

Structure of a Complete Mediator-RNA Polymerase II Pre-Initiation Complex
- PMID: 27610567
- PMCID: PMC5589196
- DOI: 10.1016/j.cell.2016.08.050
Structure of a Complete Mediator-RNA Polymerase II Pre-Initiation Complex
Abstract
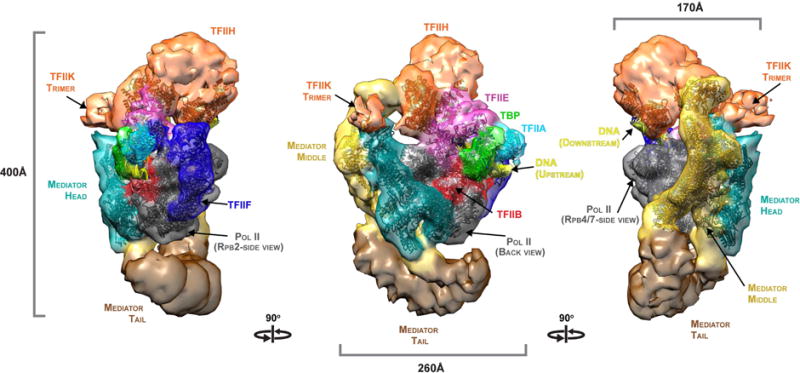
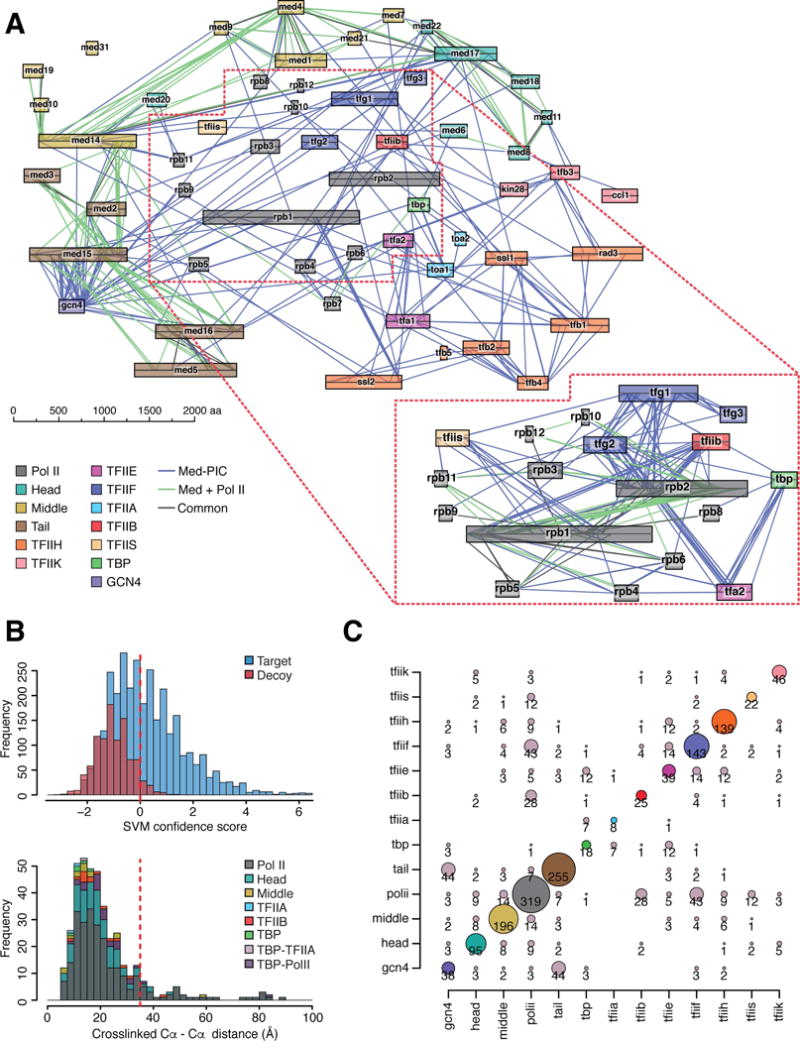
A complete, 52-protein, 2.5 million dalton, Mediator-RNA polymerase II pre-initiation complex (Med-PIC) was assembled and analyzed by cryo-electron microscopy and by chemical cross-linking and mass spectrometry. The resulting complete Med-PIC structure reveals two components of functional significance, absent from previous structures, a protein kinase complex and the Mediator-activator interaction region. It thereby shows how the kinase and its target, the C-terminal domain of the polymerase, control Med-PIC interaction and transcription.
Keywords: Mediator complex; Pre-initiation complex; RNA polymerase II carboxy-terminal domain; TFIIH; TFIIK; Transcription; cross-linking; cryo-EM; mass spectrometry.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures





References
-
- Asturias FJ, Jiang YW, Myers LC, Gustafsson CM, Kornberg RD. Conserved structures of mediator and RNA polymerase II holoenzyme. Science. 1999;283:985–987. - PubMed
-
- Baumli S, Hoeppner S, Cramer P. A conserved mediator hinge revealed in the structure of the MED7.MED21 (Med7.Srb7) heterodimer. The Journal of biological chemistry. 2005;280:18171–18178. - PubMed
-
- Beve J, Hu GZ, Myers LC, Balciunas D, Werngren O, Hultenby K, Wibom R, Ronne H, Gustafsson CM. The structural and functional role of Med5 in the yeast Mediator tail module. The Journal of biological chemistry. 2005;280:41366–41372. - PubMed
-
- Bich C, Maedler S, Chiesa K, DeGiacomo F, Bogliotti N, Zenobi R. Reactivity and applications of new amine reactive cross-linkers for mass spectrometric detection of protein-protein complexes. Analytical chemistry. 2010;82:172–179. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
