

mSWI/SNF (BAF) Complexes Are Indispensable for the Neurogenesis and Development of Embryonic Olfactory Epithelium
- PMID: 27611684
- PMCID: PMC5017785
- DOI: 10.1371/journal.pgen.1006274
mSWI/SNF (BAF) Complexes Are Indispensable for the Neurogenesis and Development of Embryonic Olfactory Epithelium
Abstract
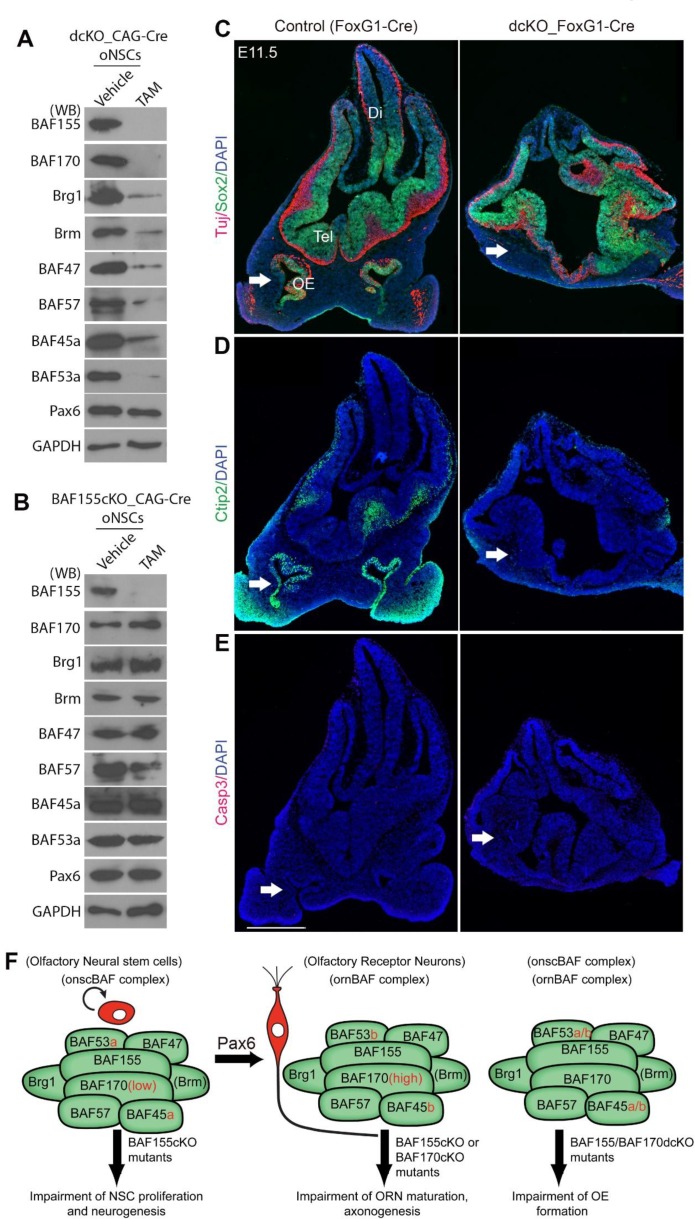
Neurogenesis is a key developmental event through which neurons are generated from neural stem/progenitor cells. Chromatin remodeling BAF (mSWI/SNF) complexes have been reported to play essential roles in the neurogenesis of the central nervous system. However, whether BAF complexes are required for neuron generation in the olfactory system is unknown. Here, we identified onscBAF and ornBAF complexes, which are specifically present in olfactory neural stem cells (oNSCs) and olfactory receptor neurons (ORNs), respectively. We demonstrated that BAF155 subunit is highly expressed in both oNSCs and ORNs, whereas high expression of BAF170 subunit is observed only in ORNs. We report that conditional deletion of BAF155, a core subunit in both onscBAF and ornBAF complexes, causes impaired proliferation of oNSCs as well as defective maturation and axonogenesis of ORNs in the developing olfactory epithelium (OE), while the high expression of BAF170 is important for maturation of ORNs. Interestingly, in the absence of BAF complexes in BAF155/BAF170 double-conditional knockout mice (dcKO), OE is not specified. Mechanistically, BAF complex is required for normal activation of Pax6-dependent transcriptional activity in stem cells/progenitors of the OE. Our findings unveil a novel mechanism mediated by the mSWI/SNF complex in OE neurogenesis and development.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures








References
-
- Treloar HB, Miller AM, Ray A, Greer CA (2010) Development of the Olfactory System. In: Menini A, editor. The Neurobiology of Olfaction. Boca Raton (FL). - PubMed
-
- Buck LB (2005) Unraveling the sense of smell (Nobel lecture). Angewandte Chemie 44: 6128–6140. - PubMed
-
- Cau E, Casarosa S, Guillemot F (2002) Mash1 and Ngn1 control distinct steps of determination and differentiation in the olfactory sensory neuron lineage. Development 129: 1871–1880. - PubMed
-
- Cau E, Gradwohl G, Fode C, Guillemot F (1997) Mash1 activates a cascade of bHLH regulators in olfactory neuron progenitors. Development 124: 1611–1621. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
