

In Silico and Biochemical Characterization of Lysozyme-Like Proteins in the Rat
- PMID: 27611690
- PMCID: PMC5017655
- DOI: 10.1371/journal.pone.0161909
In Silico and Biochemical Characterization of Lysozyme-Like Proteins in the Rat
Abstract
Background: Spermatogenesis and sperm maturation in the male reproductive tract is dictated by a variety of proteins secreted in the testis and epididymis. Though the proteome of these tissues is known, the functional role of many of these proteins remains uncharacterized. In this study, we characterize the rat Lysozyme-like (Lyzl) genes and proteins.
Methods: In silico tools were used to predict the primary, secondary and tertiary structures. Reverse transcription PCR, immunofluorescence and immunoblotting were used to determine the expression pattern. Lysozyme like enzyme activity was assessed by standard assays.
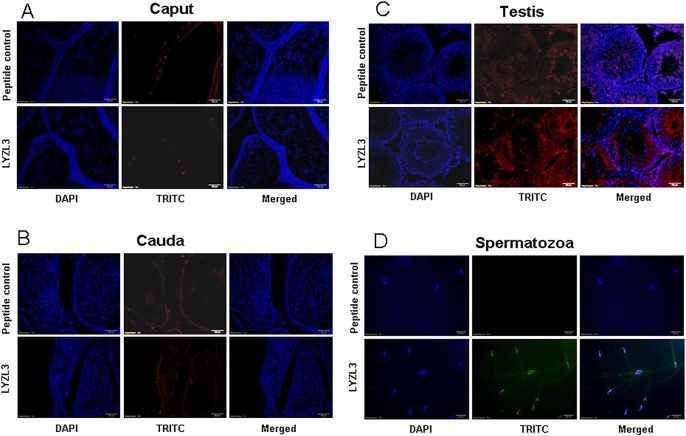
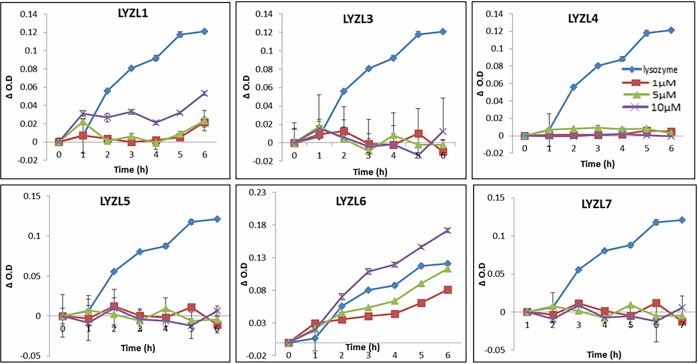
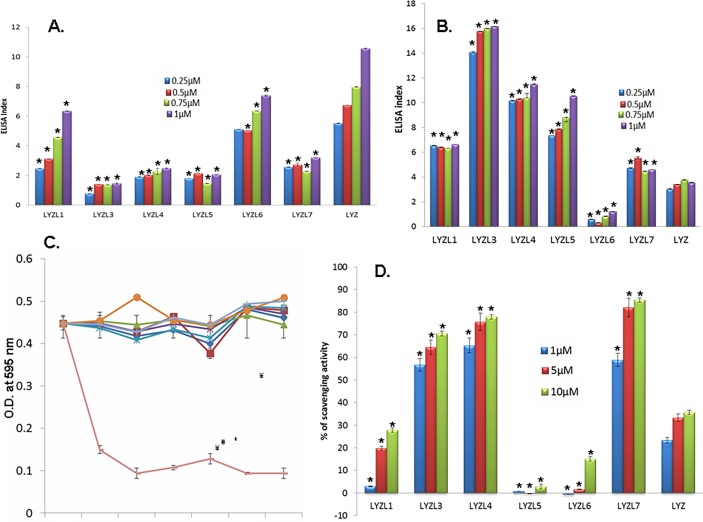
Results: Six rat Lyzl genes namely Lyzl1, Lyzl3, Lyzl4, Lyzl5, Lyzl6 and Lyzl7 were found to be highly conserved among the vertebrates with higher homology to mouse counterparts than with human counterparts. All the LYZL proteins contained the characteristic 4 disulfide bridges similar to c-type lysozyme. Only LYZL 1 and 6, conserved the active site amino acids of the lysozyme. Molecular modeling studies indicated that LYZL proteins exhibit strikingly similar three-dimensional structures among themselves. The secondary structure analysis of the recombinant LYZL proteins indicated the presence of α-helix, β-sheet and random coil with α-helix being the majority. Docking studies indicated the peptidoglycan binding nature of LYZL proteins. All the rat Lyzl mRNA transcripts (Lyzl1, Lyzl3, Lyzl4, Lyzl5, Lyzl6 and Lyzl7) are predominantly expressed in testes though some of them are expressed in tissues other than reproductive tract. Their expression was androgen independent. The rat LYZL proteins are localized in the germinal epithelium and on the spermatozoa. Recombinant LYZL1 and 6 possessed muramidase, isopeptidase and antibacterial activities. The mechanism of antibacterial action of LYZL1 and LYZL6 involved bacterial membrane damage and leakage of cellular contents. Only LYZL1 and 6 possess peptidoglycan binding ability, whereas LYZL3, LYZL4 and LYZL5 possess hyaluronan binding ability suggesting a possible functional divergence of these proteins. LYZL3, LYZL4 and LYZL7 possessed free radical scavenging property, suggesting that they may act as antioxidants.
Conclusion: The divergent properties of LYZL proteins indicate that they may have a role in sperm function, innate immunity and other physiological process as well.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
















References
-
- Kirchhoff C (1998) Molecular characterization of epididymal proteins. Reviews of reproduction 3: 86–95. - PubMed
-
- Jiborn T, Abrahamson M, Wallin H, Malm J, Lundwall A, Gadaleanu V, et al. (2004) Cystatin C is highly expressed in the human male reproductive system. Journal of andrology 25: 564–572. - PubMed
-
- Jin YZ, Bannai S, Dacheux F, Dacheux JL, Okamura N (1997) Direct evidence for the secretion of lactoferrin and its binding to sperm in the porcine epididymis. Molecular reproduction and development 47: 490–496. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
