

Role of Host-Driven Mutagenesis in Determining Genome Evolution of Sigma Virus (DMelSV; Rhabdoviridae) in Drosophila melanogaster
- PMID: 27614234
- PMCID: PMC5630973
- DOI: 10.1093/gbe/evw212
Role of Host-Driven Mutagenesis in Determining Genome Evolution of Sigma Virus (DMelSV; Rhabdoviridae) in Drosophila melanogaster
Abstract
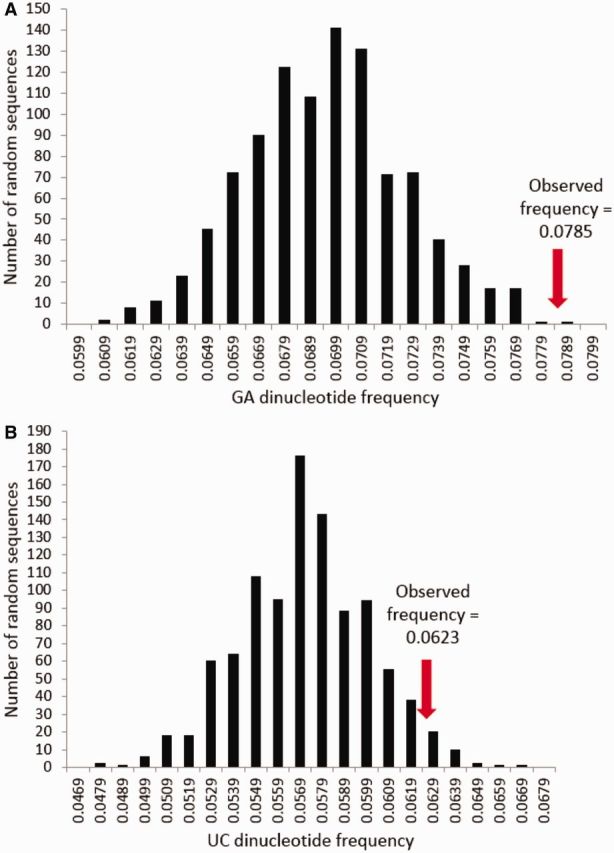
Sigma virus (DMelSV) is ubiquitous in natural populations of Drosophila melanogaster. Host-mediated, selective RNA editing of adenosines to inosines (ADAR) may contribute to control of viral infection by preventing transcripts from being transported into the cytoplasm or being translated accurately; or by increasing the viral genomic mutation rate. Previous PCR-based studies showed that ADAR mutations occur in DMelSV at low frequency. Here we use SOLiDTM deep sequencing of flies from a single host population from Athens, GA, USA to comprehensively evaluate patterns of sequence variation in DMelSV with respect to ADAR. GA dinucleotides, which are weak targets of ADAR, are strongly overrepresented in the positive strand of the virus, consistent with selection to generate ADAR resistance on this complement of the transient, double-stranded RNA intermediate in replication and transcription. Potential ADAR sites in a worldwide sample of viruses are more likely to be "resistant" if the sites do not vary among samples. Either variable sites are less constrained and hence are subject to weaker selection than conserved sites, or the variation is driven by ADAR. We also find evidence of mutations segregating within hosts, hereafter referred to as hypervariable sites. Some of these sites were variable only in one or two flies (i.e., rare); others were shared by four or even all five of the flies (i.e., common). Rare and common hypervariable sites were indistinguishable with respect to susceptibility to ADAR; however, polymorphism in rare sites were more likely to be consistent with the action of ADAR than in common ones, again suggesting that ADAR is deleterious to the virus. Thus, in DMelSV, host mutagenesis is constraining viral evolution both within and between hosts.
Keywords: ADAR; RNA virus; immunity; mutation; phylogeny; quasispecies.
© The Author 2016. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures



References
-
- Brun G. 1963. Édude d'une association du virus et son hôte la Drosophilé: l'état stabilisé. Thèse Paris XI, Orsay, 1963.
-
- Brun G, Plus N. 1980. The viruses of Drosophila In: Ashburner M, Wright T. R. F, editors. The genetics and biology of Drosophila. London: Academic Press. p; 625–702.
-
- Carpenter JA, Obbard DJ, Maside X, Jiggins FM. 2007. The recent spread of a vertically transmitted virus through populations of Drosophila melanogaster. Mol Ecol. 16:3947–3954. doi: 10.1111/j.1365-294X.2007.03460.x - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
