

Ontogenic Changes and Differential Localization of T-type Ca(2+) Channel Subunits Cav3.1 and Cav3.2 in Mouse Hippocampus and Cerebellum
- PMID: 27616982
- PMCID: PMC4999439
- DOI: 10.3389/fnana.2016.00083
Ontogenic Changes and Differential Localization of T-type Ca(2+) Channel Subunits Cav3.1 and Cav3.2 in Mouse Hippocampus and Cerebellum
Abstract
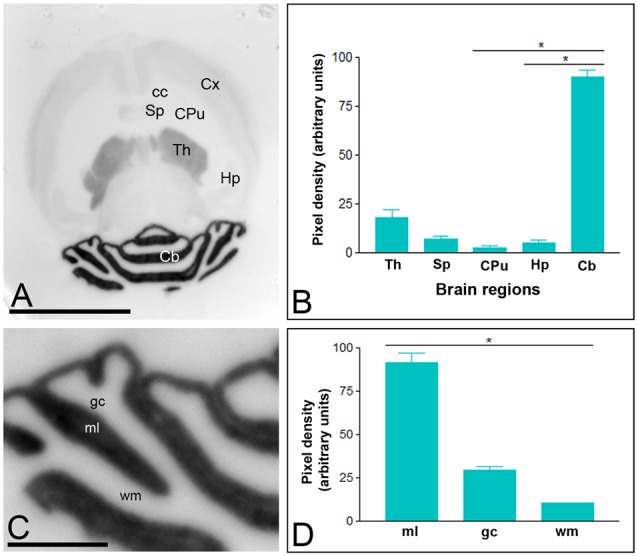
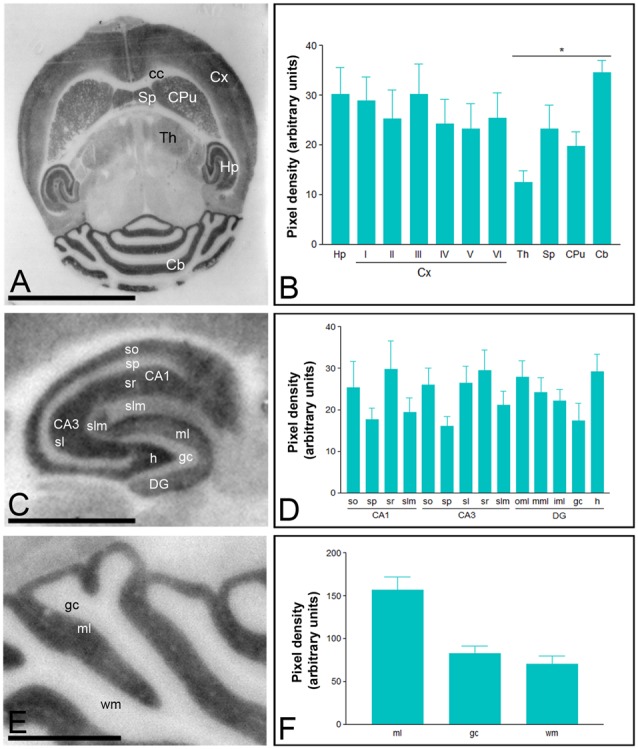
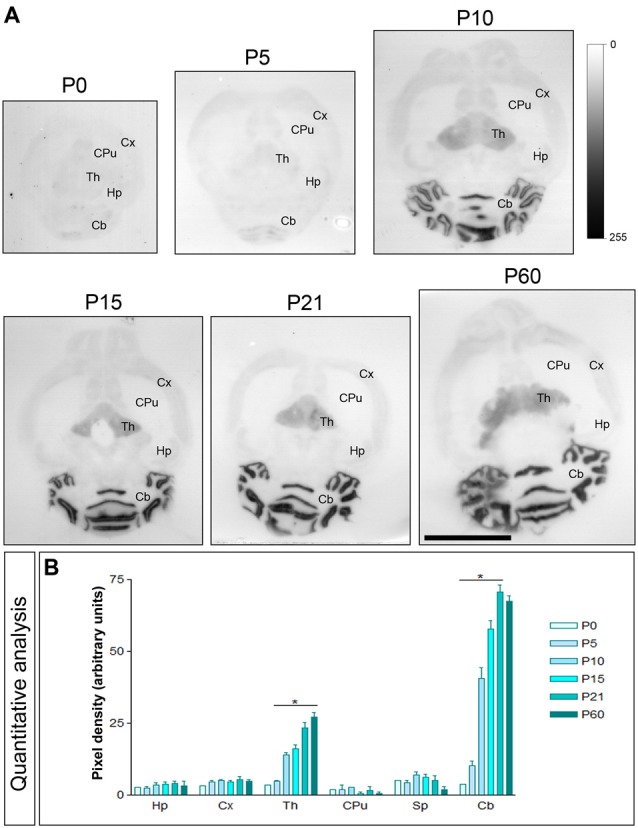
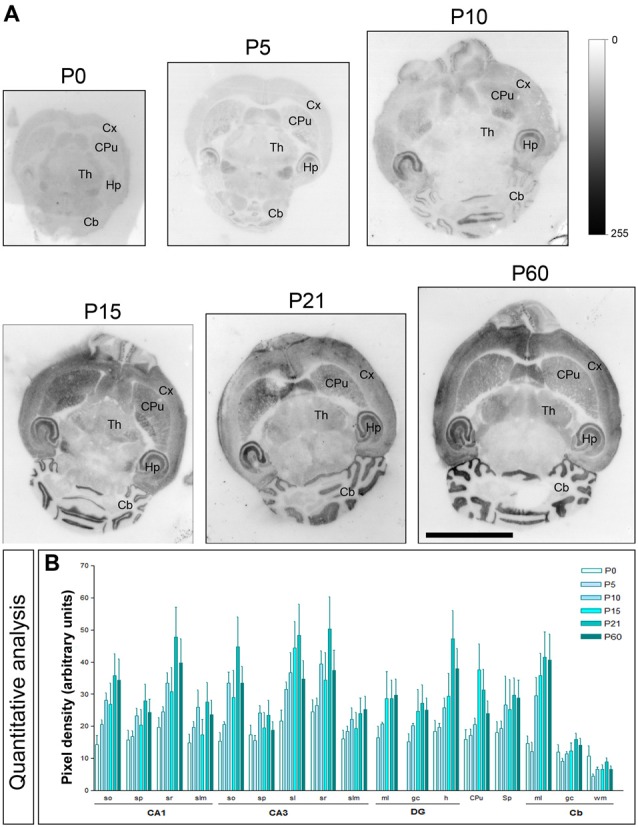
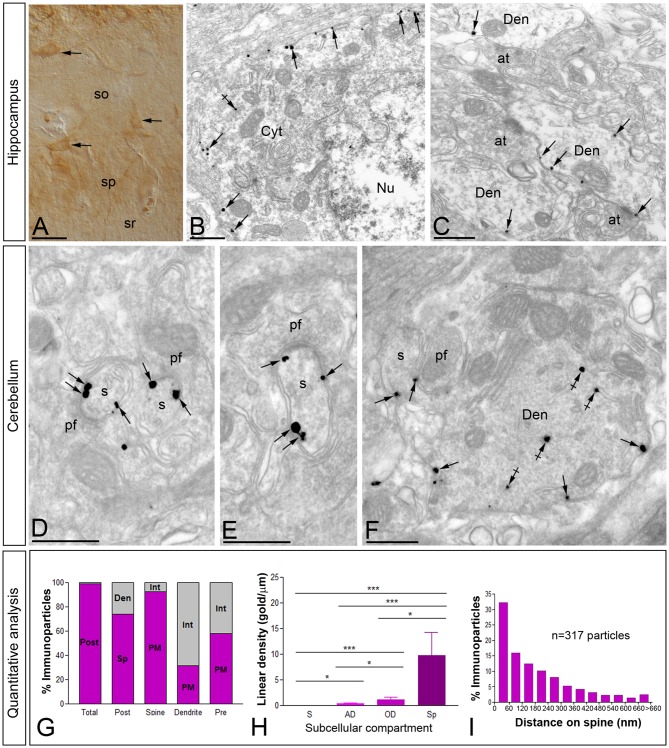
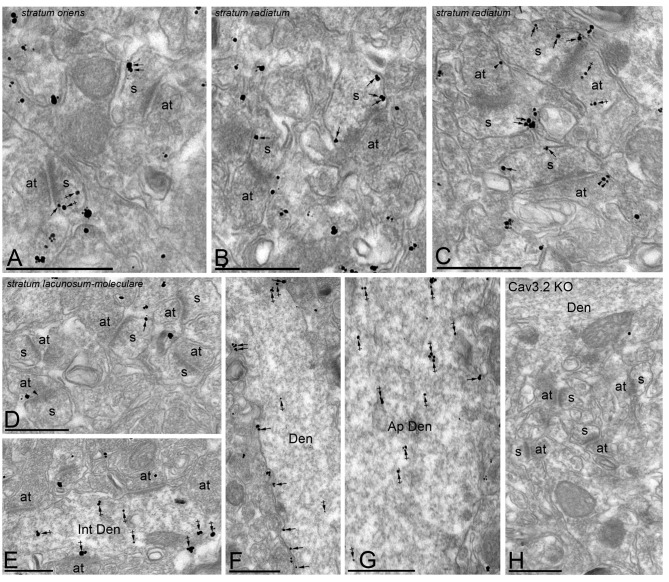
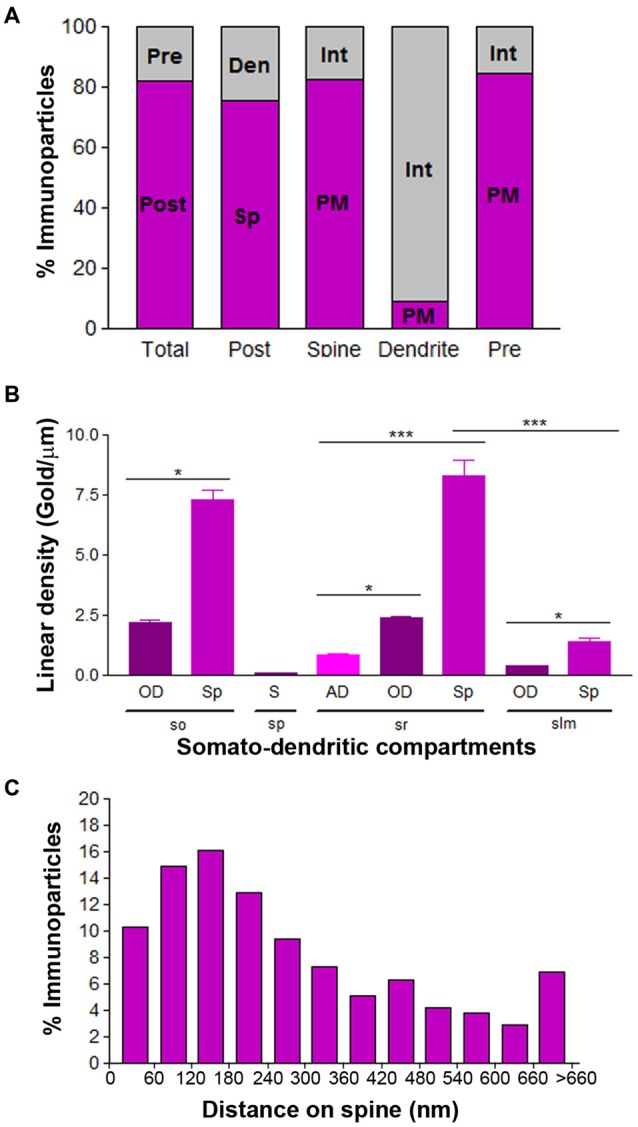
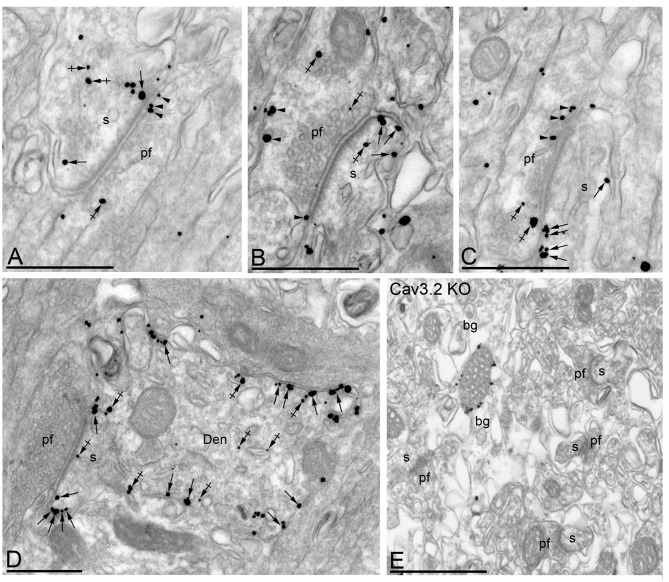
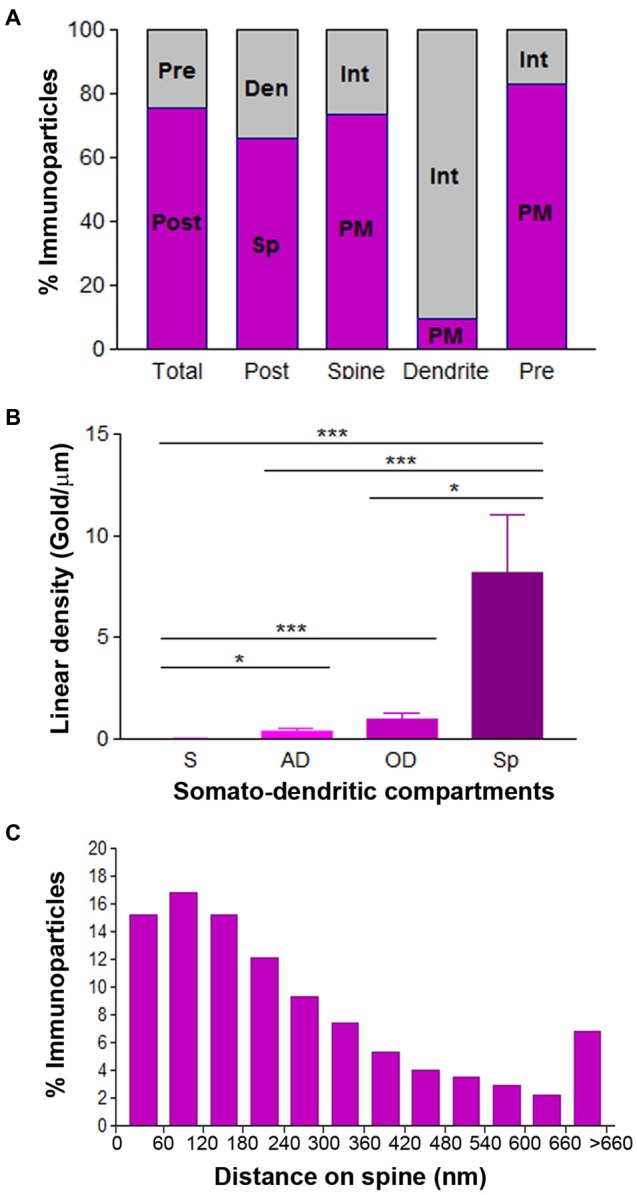
T-type calcium (Ca(2+)) channels play a central role in regulating membrane excitability in the brain. Although the contributions of T-type current to neuron output is often proposed to reflect a differential distribution of T-type channel subtypes to somato-dendritic compartments, their precise subcellular distributions in central neurons are not fully determined. Using histoblot and high-resolution immunoelectron microscopic techniques, we have investigated the expression, regional distribution and subcellular localization of T-type Cav3.1 and Cav3.2 channel subunits in the adult brain, as well as the ontogeny of expression during postnatal development. Histoblot analysis showed that Cav3.1 and Cav3.2 proteins were widely expressed in the brain, with mostly non-overlapping patterns. Cav3.1 showed the highest expression level in the molecular layer (ml) of the cerebellum (Cb), and Cav3.2 in the hippocampus (Hp) and the ml of Cb. During development, levels of Cav3.1 and Cav3.2 increased with age, although there were marked region- and developmental stage-specific differences in their expression. At the cellular and subcellular level, immunoelectron microscopy showed that labeling for Cav3.1 was present in somato-dendritic domains of hippocampal interneurons and Purkinje cells (PCs), while Cav3.2 was present in somato-dendritic domains of CA1 pyramidal cells, hippocampal interneurons and PCs. Most of the immunoparticles for Cav3.1 and Cav3.2 were either associated with the plasma membrane or the intracellular membranes, with notable differences depending on the compartment. Thus, Cav3.1 was mainly located in the plasma membrane of interneurons, whereas Cav3.2 was mainly located in the plasma membrane of dendritic spines and had a major intracellular distribution in dendritic shafts. In PCs, Cav3.1 and Cav3.2 showed similar distribution patterns. In addition to its main postsynaptic distribution, Cav3.2 but not Cav3.1 was also detected in axon terminals establishing excitatory synapses. These results shed new light on the subcellular localization of T-type channel subunits and provide evidence for the non-uniform distribution of Cav3.1 and Cav3.2 subunits over the plasma membrane of central neurons, which may account for the functional heterogeneity of T-type mediated current.
Keywords: calcium channels; cerebellum; electron microscopy; hippocampus; immunohistochemistry; quantification.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous