

L1-associated genomic regions are deleted in somatic cells of the healthy human brain
- PMID: 27618310
- PMCID: PMC5127747
- DOI: 10.1038/nn.4388
L1-associated genomic regions are deleted in somatic cells of the healthy human brain
Erratum in
-
Corrigendum: L1-associated genomic regions are deleted in somatic cells of the healthy human brain.Nat Neurosci. 2017 Sep 28;20(10):1427. doi: 10.1038/nn1017-1427a. Nat Neurosci. 2017. PMID: 28949329 No abstract available.
-
Author Correction: L1-associated genomic regions are deleted in somatic cells of the healthy human brain.Nat Neurosci. 2018 Jul;21(7):1016. doi: 10.1038/s41593-018-0131-3. Nat Neurosci. 2018. PMID: 29703932
Abstract
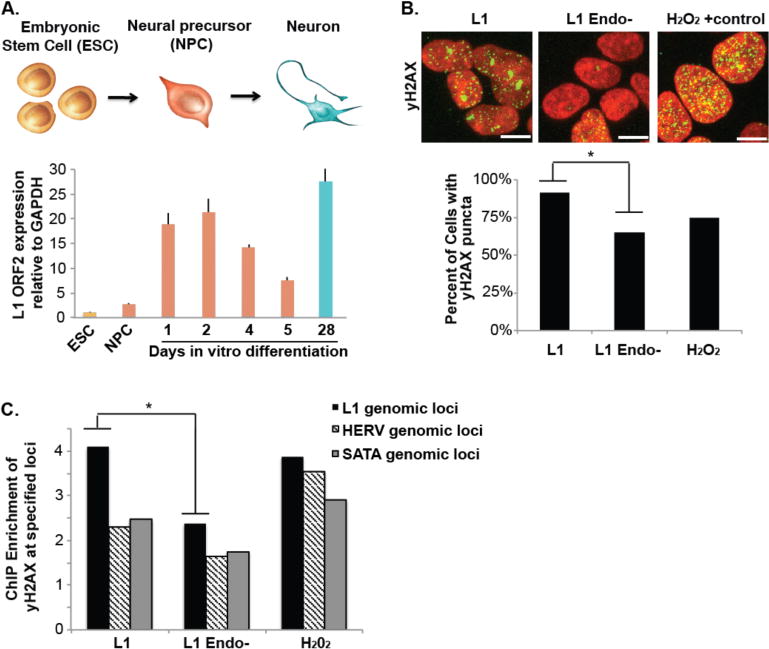
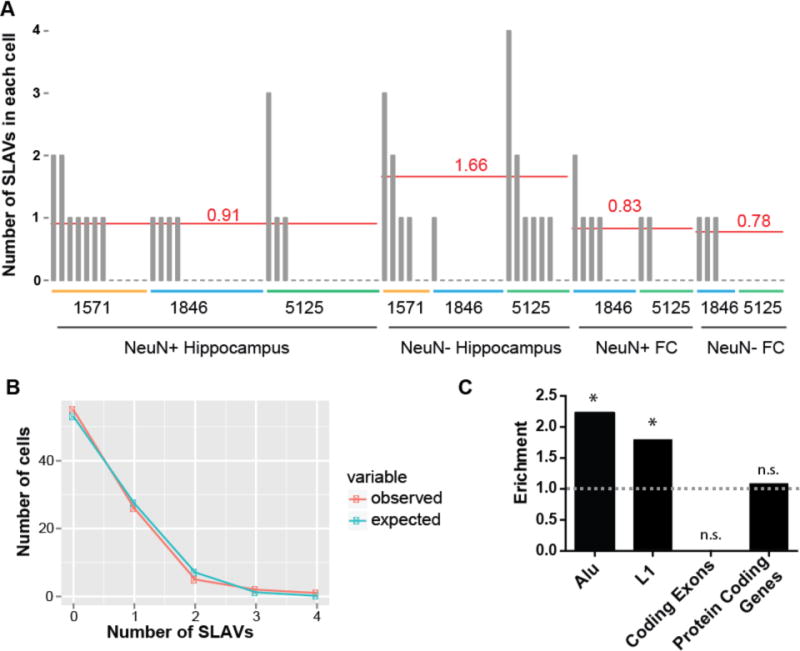
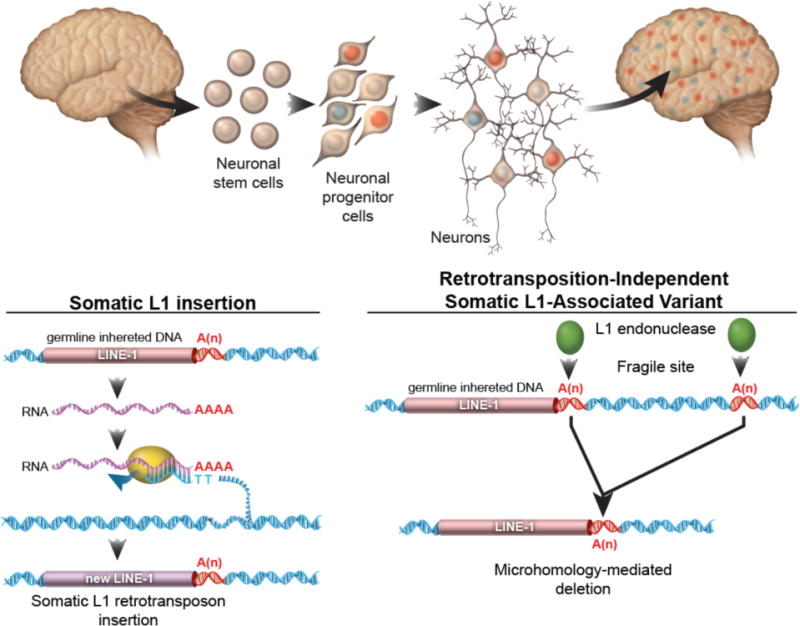
The healthy human brain is a mosaic of varied genomes. Long interspersed element-1 (LINE-1 or L1) retrotransposition is known to create mosaicism by inserting L1 sequences into new locations of somatic cell genomes. Using a machine learning-based, single-cell sequencing approach, we discovered that somatic L1-associated variants (SLAVs) are composed of two classes: L1 retrotransposition insertions and retrotransposition-independent L1-associated variants. We demonstrate that a subset of SLAVs comprises somatic deletions generated by L1 endonuclease cutting activity. Retrotransposition-independent rearrangements in inherited L1s resulted in the deletion of proximal genomic regions. These rearrangements were resolved by microhomology-mediated repair, which suggests that L1-associated genomic regions are hotspots for somatic copy number variants in the brain and therefore a heritable genetic contributor to somatic mosaicism. We demonstrate that SLAVs are present in crucial neural genes, such as DLG2 (also called PSD93), and affect 44-63% of cells of the cells in the healthy brain.
Figures
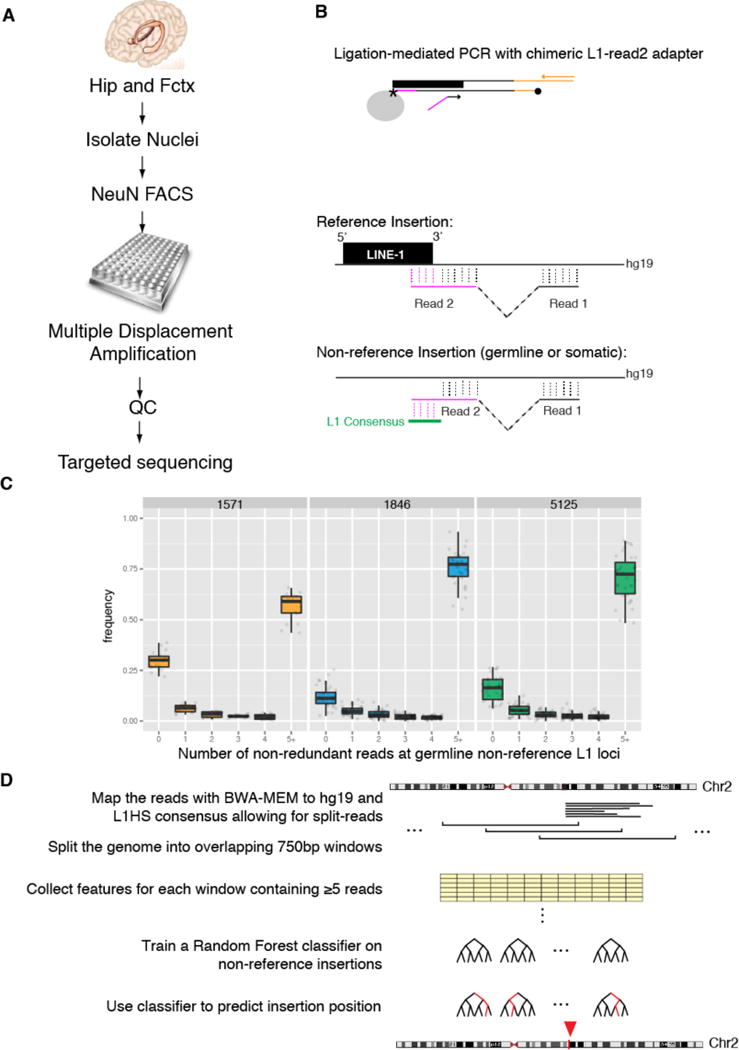
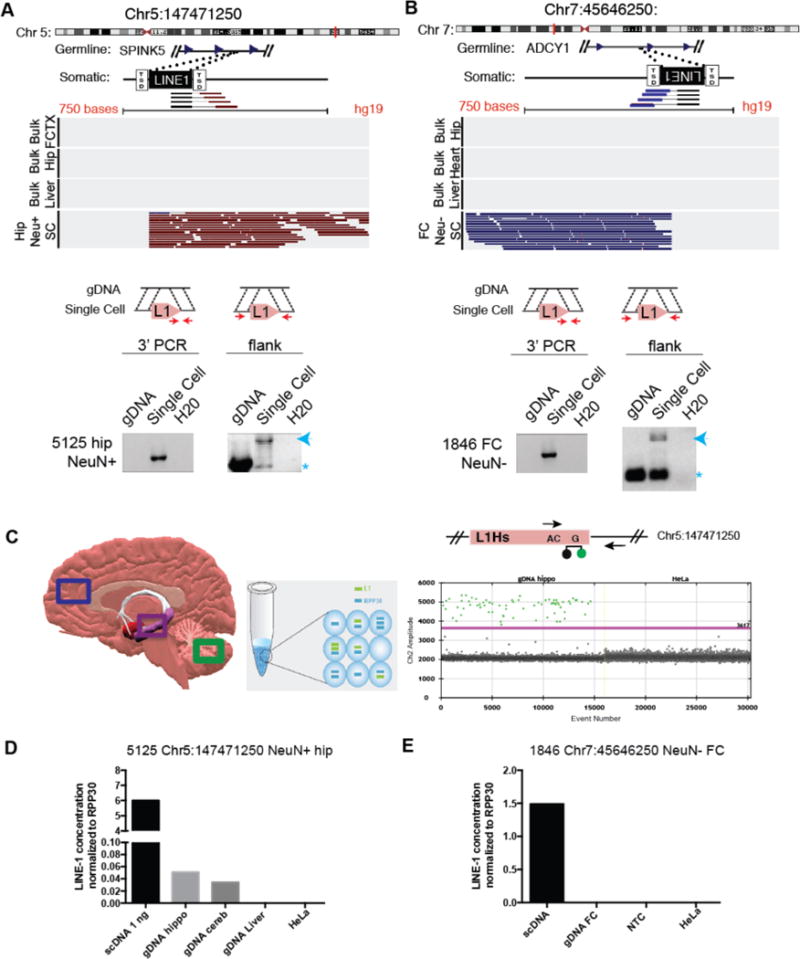
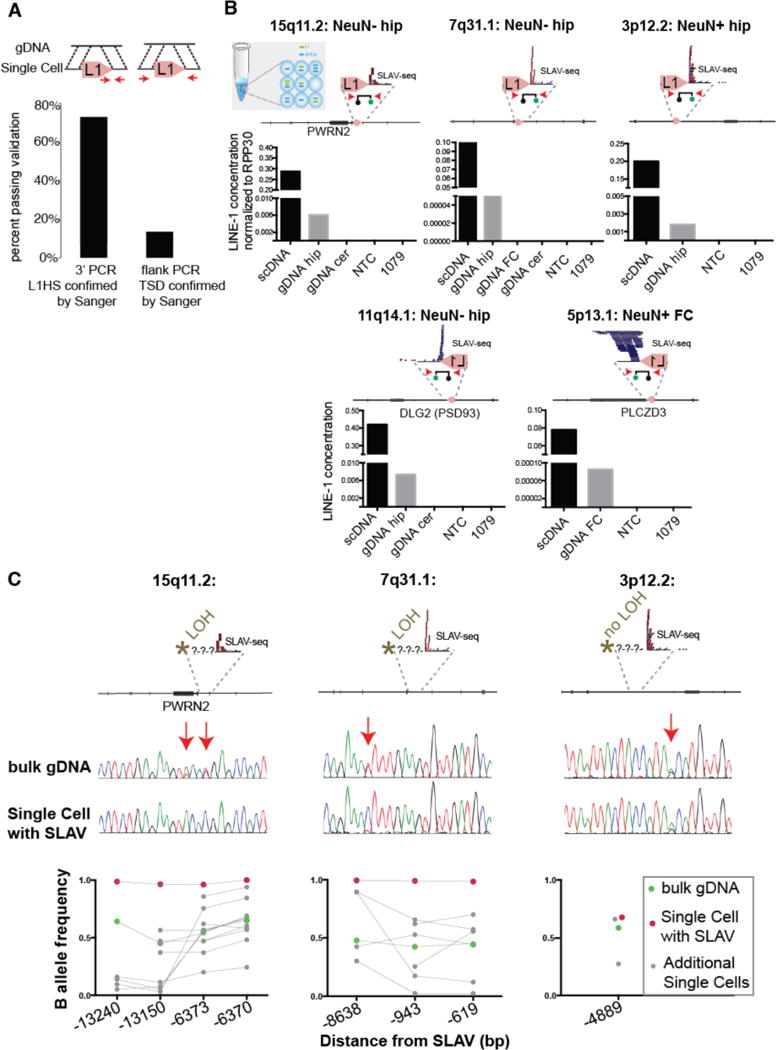
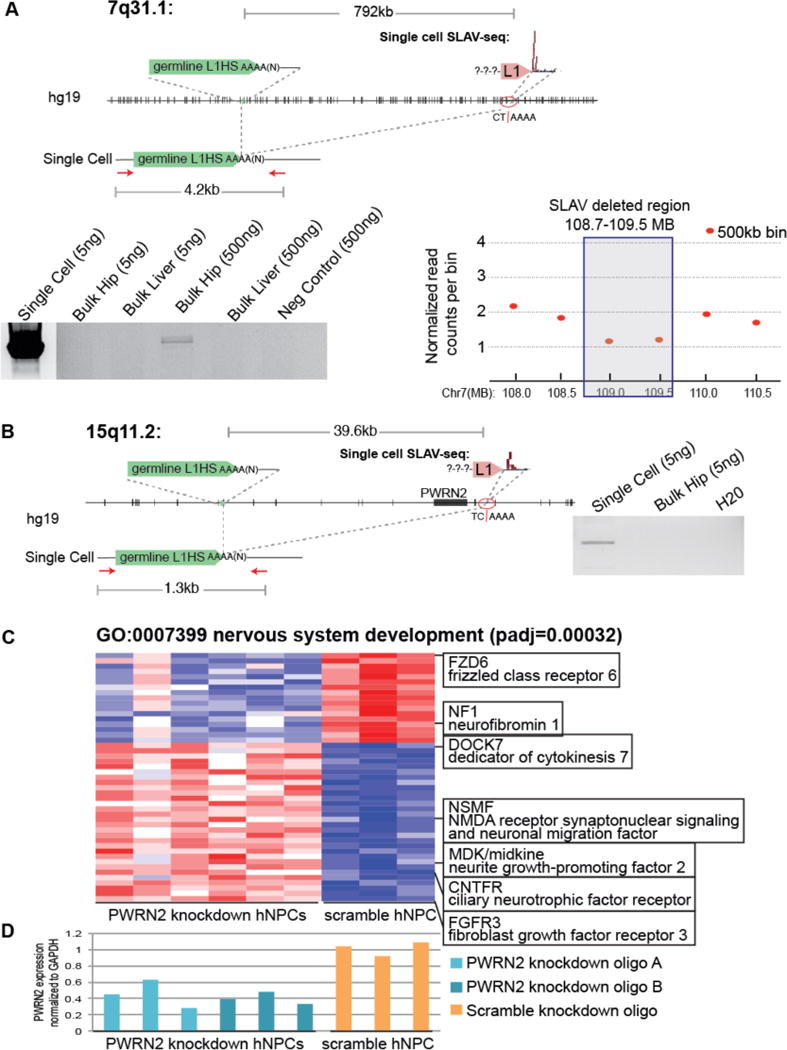
References
-
- Muotri AR, et al. Somatic mosaicism in neuronal precursor cells mediated by L1 retrotransposition. Nature. 2005;435:903–910. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
