

Mitogen-Activated Protein Kinase Cascade MKK7-MPK6 Plays Important Roles in Plant Development and Regulates Shoot Branching by Phosphorylating PIN1 in Arabidopsis
- PMID: 27618482
- PMCID: PMC5019414
- DOI: 10.1371/journal.pbio.1002550
Mitogen-Activated Protein Kinase Cascade MKK7-MPK6 Plays Important Roles in Plant Development and Regulates Shoot Branching by Phosphorylating PIN1 in Arabidopsis
Abstract
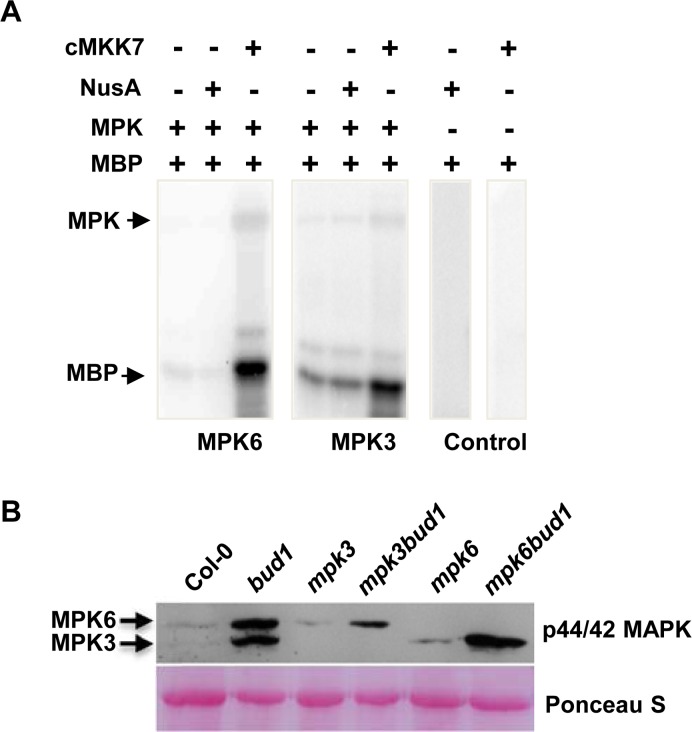
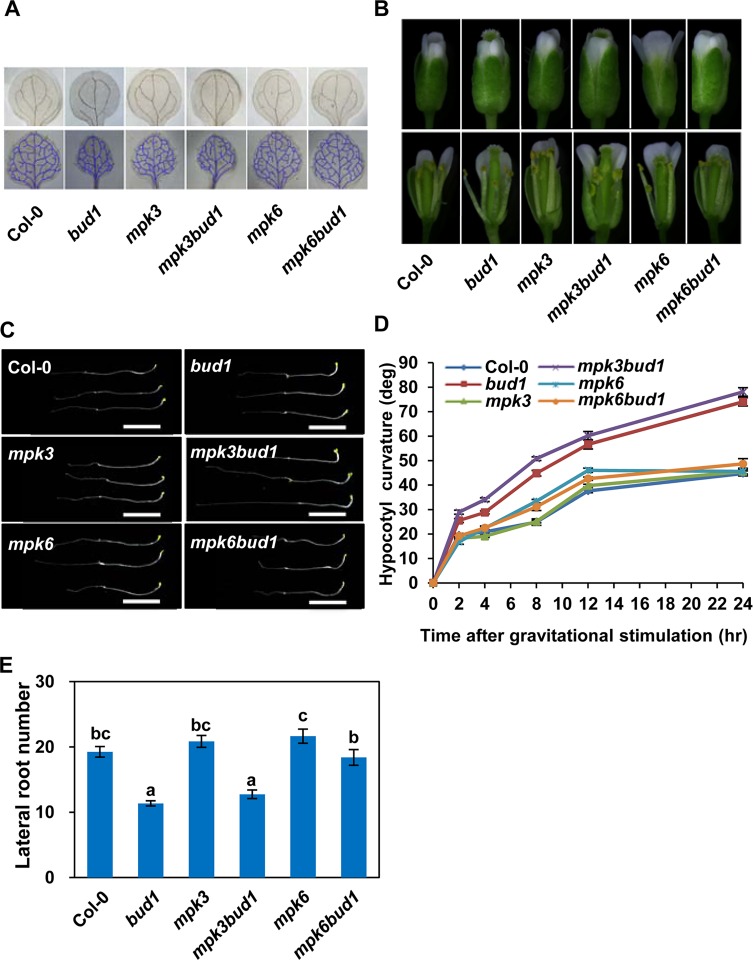
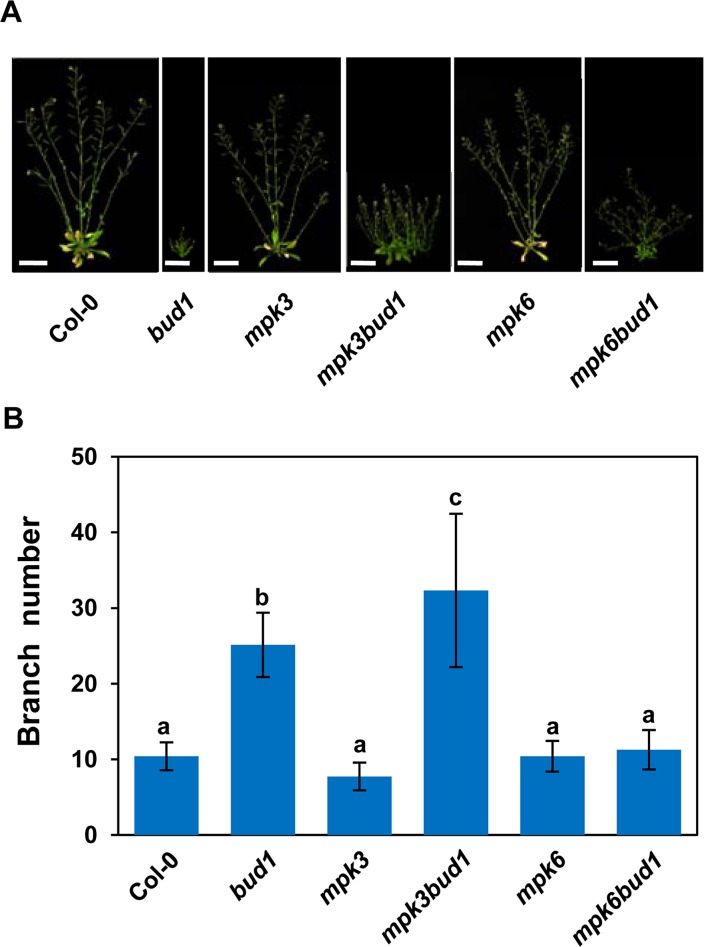
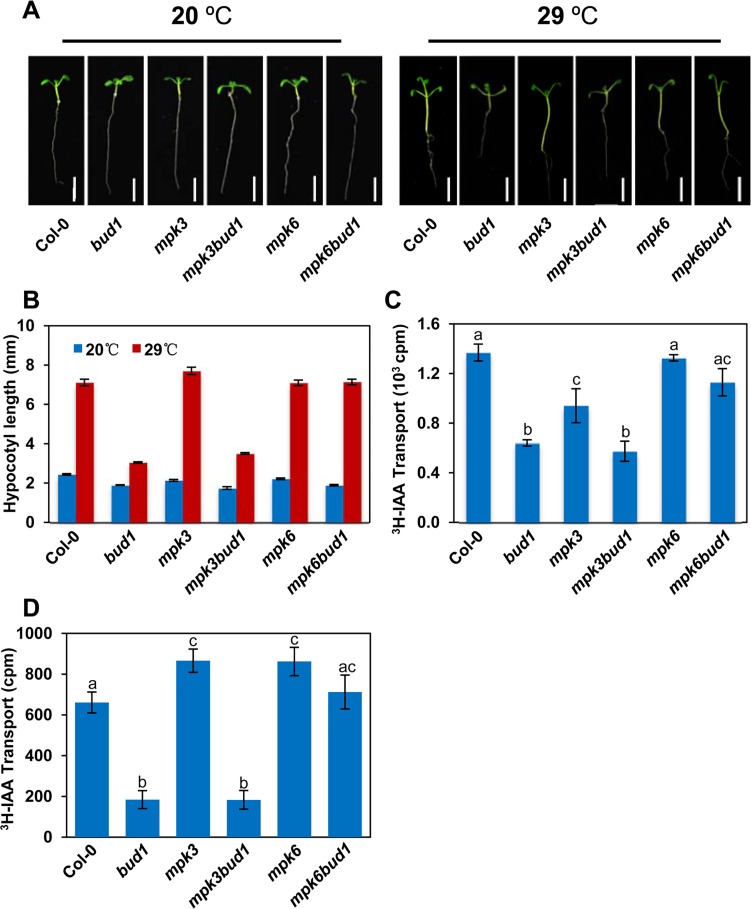
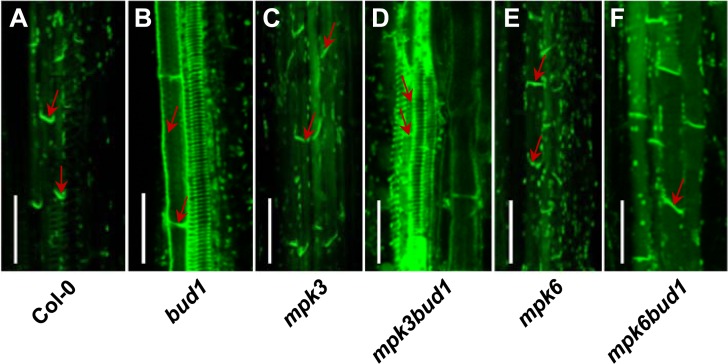
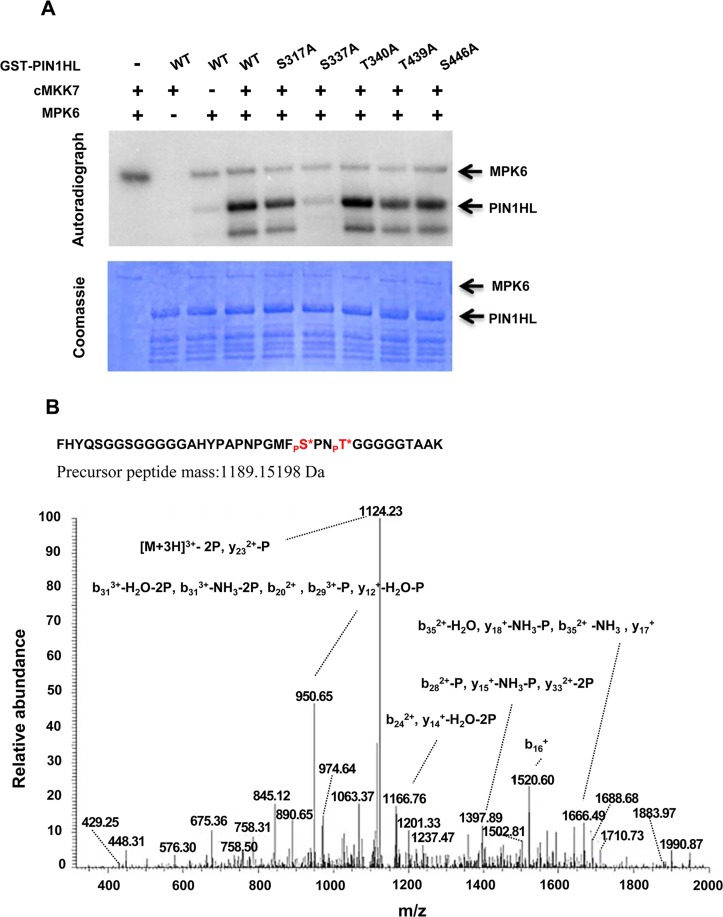
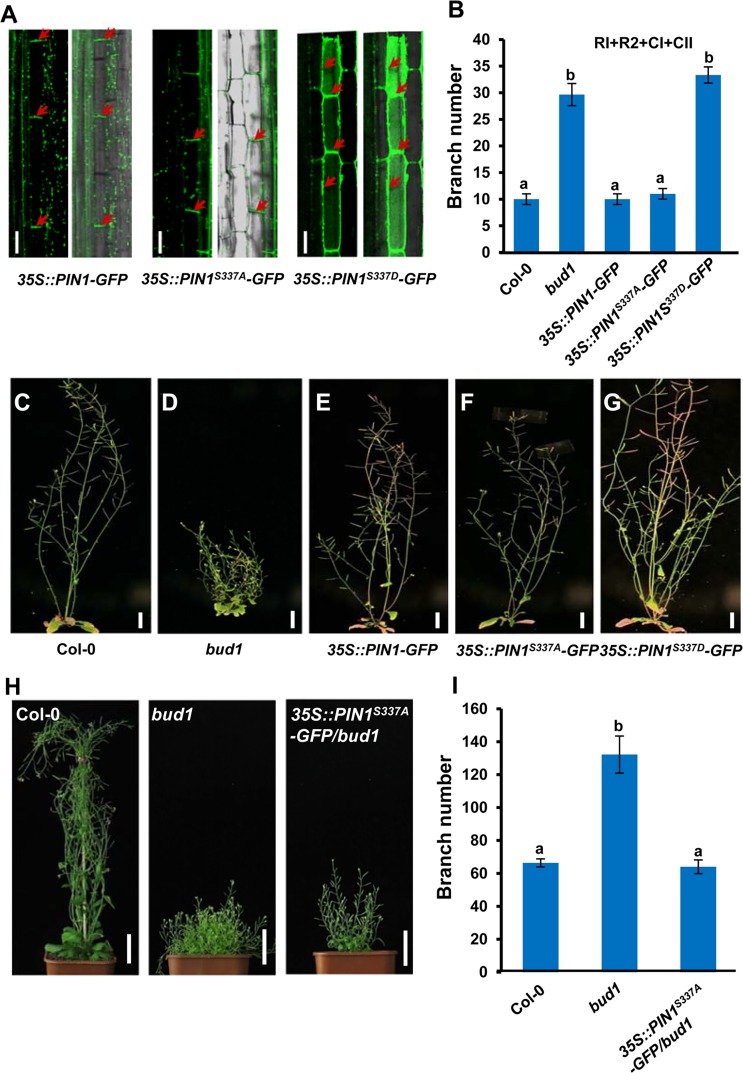
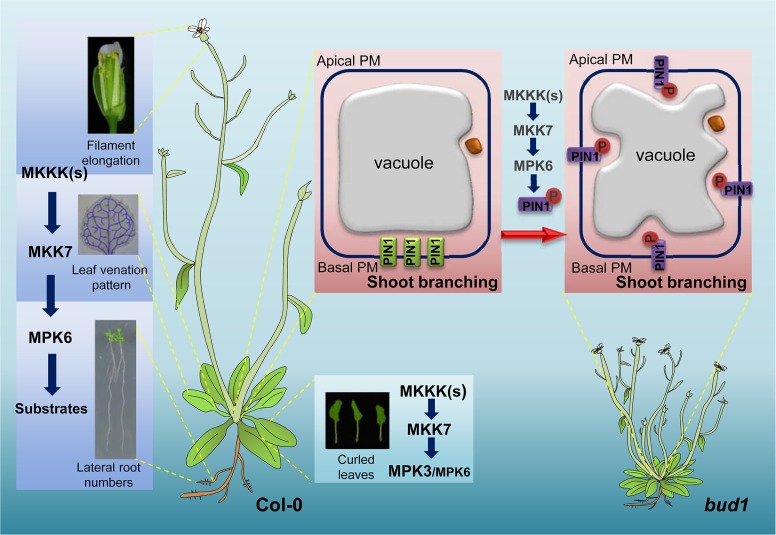
Emerging evidences exhibit that mitogen-activated protein kinase (MAPK/MPK) signaling pathways are connected with many aspects of plant development. The complexity of MAPK cascades raises challenges not only to identify the MAPK module in planta but also to define the specific role of an individual module. So far, our knowledge of MAPK signaling has been largely restricted to a small subset of MAPK cascades. Our previous study has characterized an Arabidopsis bushy and dwarf1 (bud1) mutant, in which the MAP Kinase Kinase 7 (MKK7) was constitutively activated, resulting in multiple phenotypic alterations. In this study, we found that MPK3 and MPK6 are the substrates for phosphorylation by MKK7 in planta. Genetic analysis showed that MKK7-MPK6 cascade is specifically responsible for the regulation of shoot branching, hypocotyl gravitropism, filament elongation, and lateral root formation, while MKK7-MPK3 cascade is mainly involved in leaf morphology. We further demonstrated that the MKK7-MPK6 cascade controls shoot branching by phosphorylating Ser 337 on PIN1, which affects the basal localization of PIN1 in xylem parenchyma cells and polar auxin transport in the primary stem. Our results not only specify the functions of the MKK7-MPK6 cascade but also reveal a novel mechanism for PIN1 phosphorylation, establishing a molecular link between the MAPK cascade and auxin-regulated plant development.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Group M. Mitogen-activated protein kinase cascades in plants: a new nomenclature. Trends Plant Sci. 2002;7(7):301–8. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
