

Denervation drives mitochondrial dysfunction in skeletal muscle of octogenarians
- PMID: 27619626
- PMCID: PMC5157074
- DOI: 10.1113/JP272487
Denervation drives mitochondrial dysfunction in skeletal muscle of octogenarians
Abstract
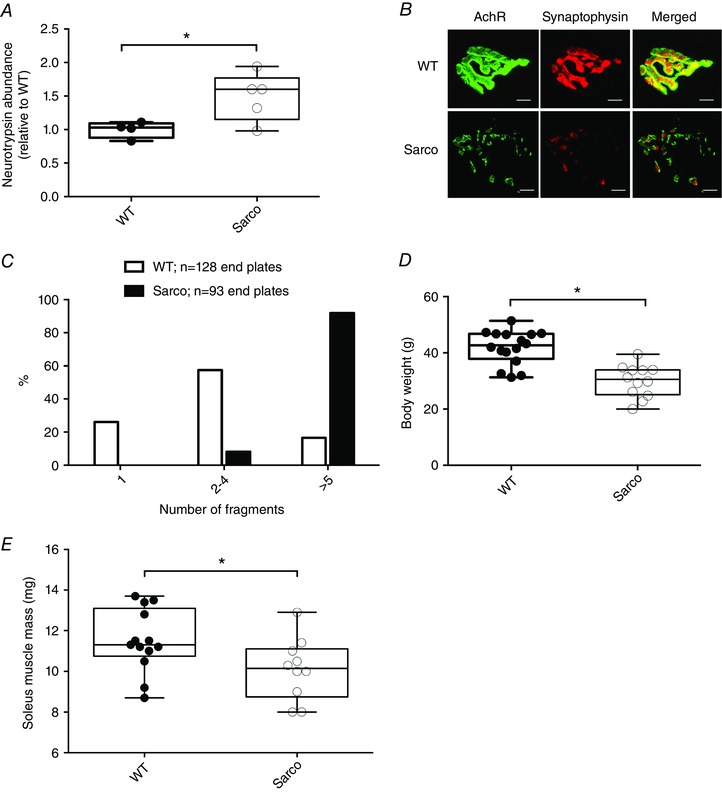
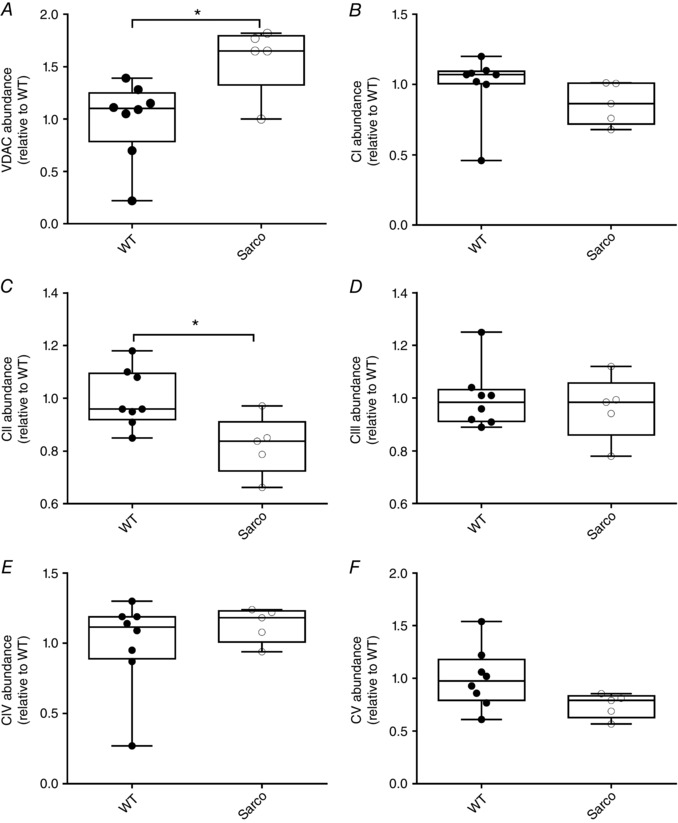
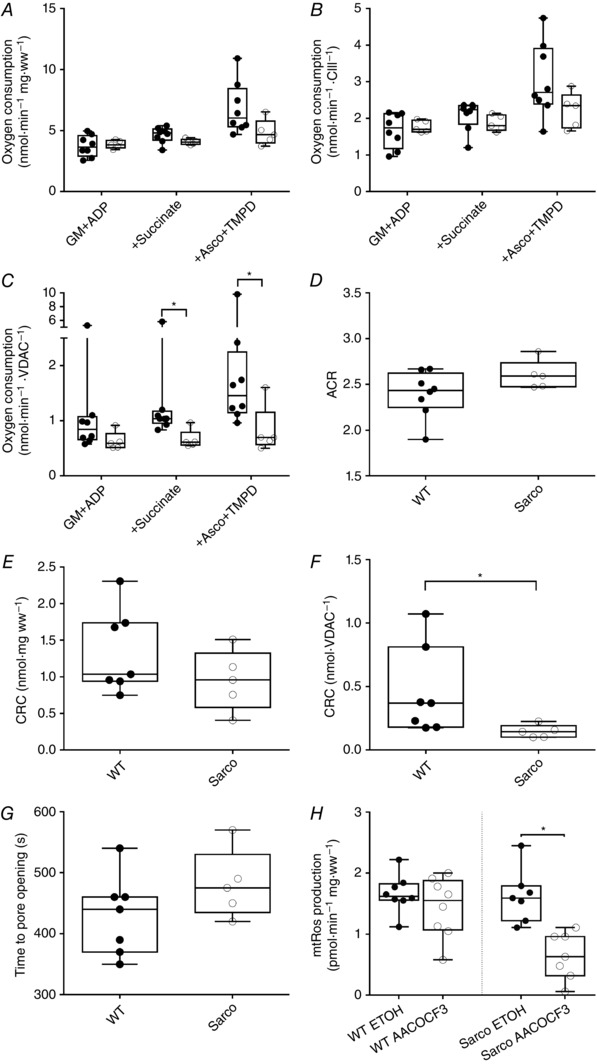
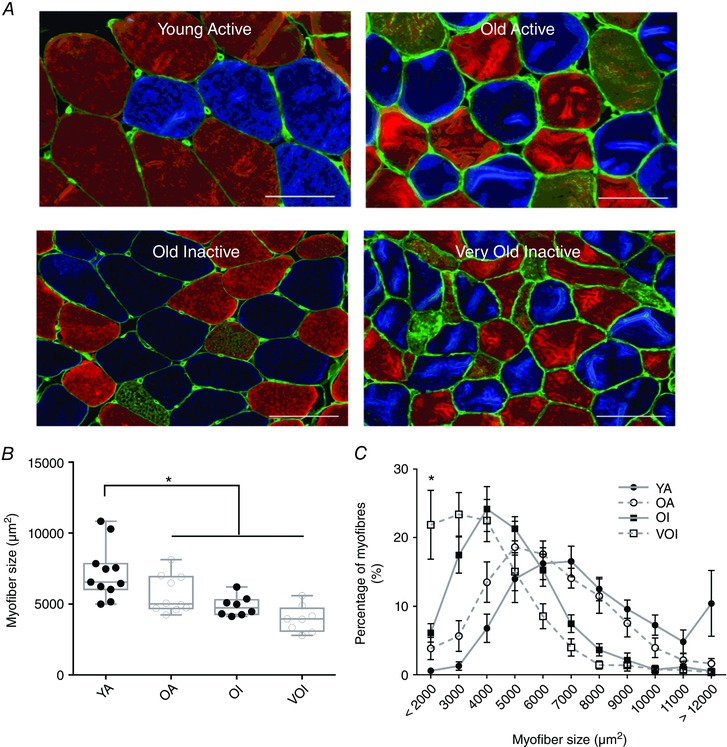
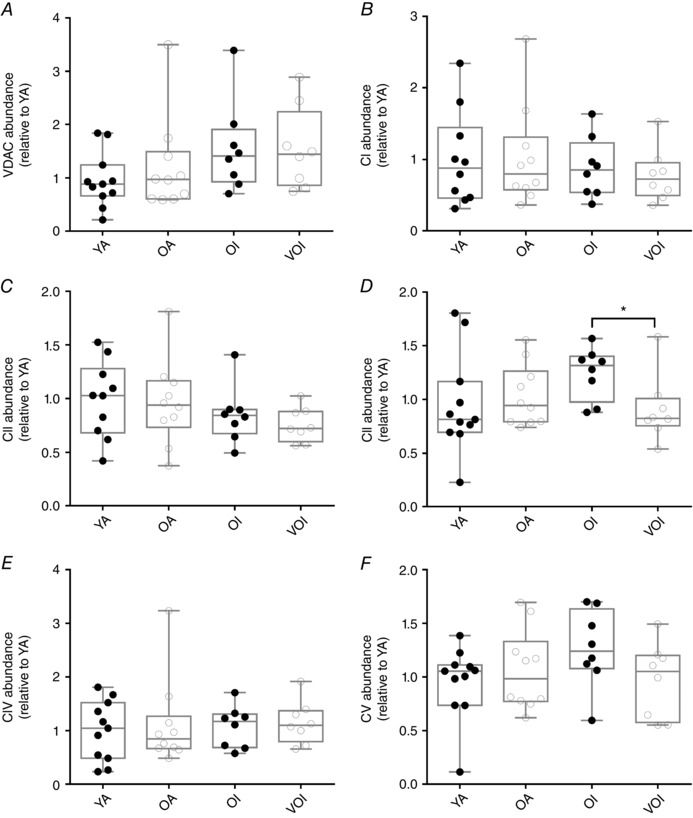
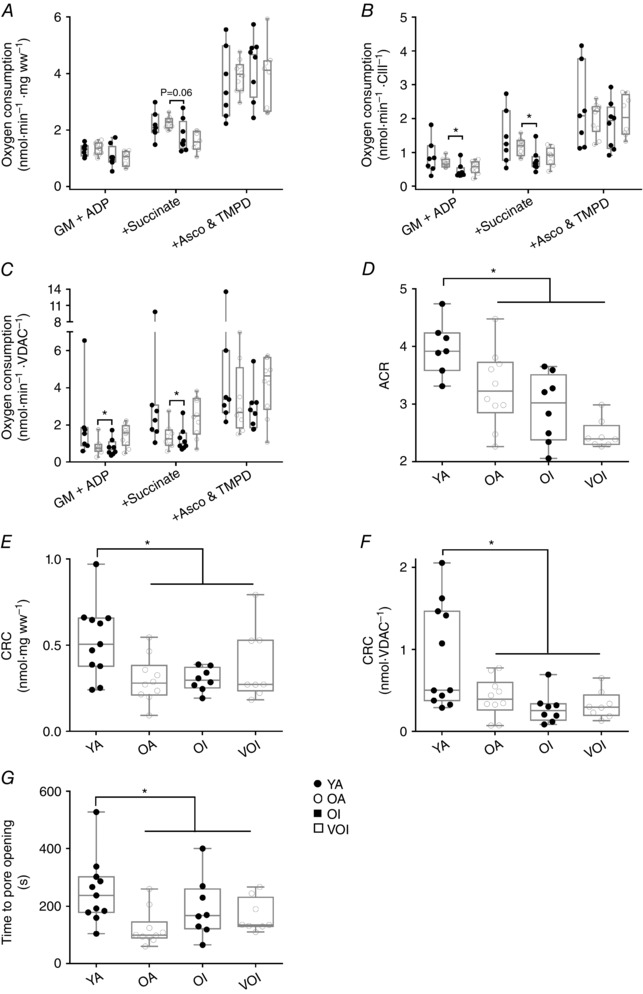
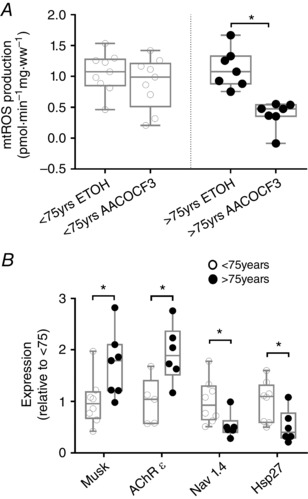
Key points: Mitochondria are frequently implicated in the ageing of skeletal muscle, although the role of denervation in modulating mitochondrial function in ageing muscle is unknown. We show that increased sensitivity to apoptosis initiation occurs prior to evidence of persistent denervation and is thus a primary mitochondrial defect in ageing muscle worthy of therapeutic targeting. However, at more advanced age, mitochondrial function changes are markedly impacted by persistent sporadic myofibre denervation, suggesting the mitochondrion may be a less viable therapeutic target.
Abstract: Experimental denervation modulates mitochondrial function, where changes in both reactive oxygen species (ROS) and sensitivity to permeability transition are implicated in the resultant muscle atrophy. Notably, although denervation occurs sporadically in ageing muscle, its impact on ageing muscle mitochondria is unknown. Because this information has important therapeutic implications concerning targeting the mitochondrion in ageing muscle, we examined mitochondrial function in skeletal muscle from four groups of humans, comprising two active (mean ± SD age: 23.7 ± 2.7 years and 71.2 ± 4.9 years) and two inactive groups (64.8 ± 3.1 years and 82.5 ± 4.8 years), and compared this with a murine model of sporadic denervation. We tested the hypothesis that, although some alterations of mitochondrial function in aged muscle are attributable to a primary organelle defect, mitochondrial dysfunction would be impacted by persistent denervation in advanced age. Both ageing in humans and sporadic denervation in mice increased mitochondrial sensitivity to permeability transition (humans, P = 0.004; mice, P = 0.01). To determine the contribution of sporadic denervation to mitochondrial function, we pharmacologically inhibited the denervation-induced ROS response. This reduced ROS emission by 60% (P = 0.02) in sporadically denervated mouse muscle, which is similar to that seen in humans older than 75 years (-66%, P = 0.02) but not those younger than 75 years. We conclude that an increased sensitivity to permeability transition is a primary mitochondrial defect in ageing muscle. However, at more advanced age, when muscle atrophy becomes more clinically severe, mitochondrial function changes are markedly impacted by persistent sporadic denervation, making the mitochondrion a less viable therapeutic target.
Keywords: aging; denervation; mitochondria; muscle atrophy; neuromuscular junction; sarcopenia.
© 2016 The Authors. The Journal of Physiology © 2016 The Physiological Society.
Figures
References
-
- Ansved T & Larsson L (1990). Effects of denervation on enzyme‐histochemical and morphometrical properties of the rat soleus muscle in relation to age. Acta Physiol Scand 139, 297–304. - PubMed
-
- Bergstrom J ( 1975). Percutaneous needle biopsy of skeletal muscle in physiological and clinical research. Scand J Clin Lab Invest 35, 609–616. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources