

Critical Role for Interleukin-25 in Host Protective Th2 Memory Response against Heligmosomoides polygyrus bakeri
- PMID: 27620722
- PMCID: PMC5116711
- DOI: 10.1128/IAI.00180-16
Critical Role for Interleukin-25 in Host Protective Th2 Memory Response against Heligmosomoides polygyrus bakeri
Abstract
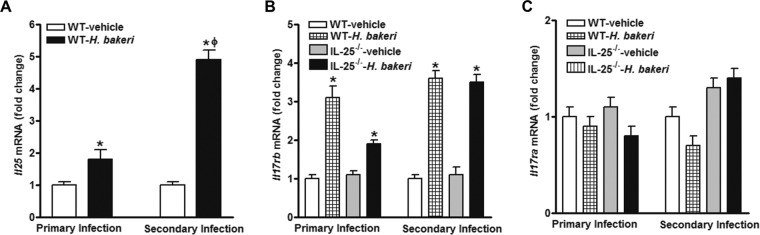
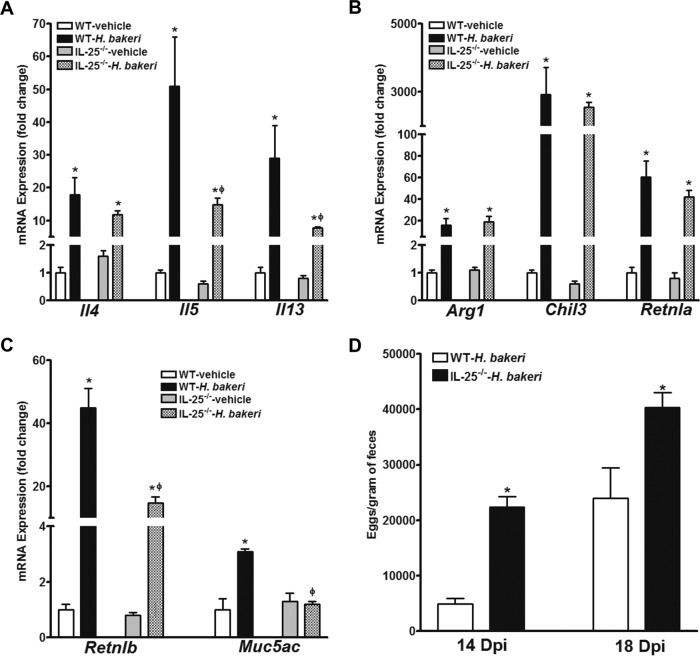
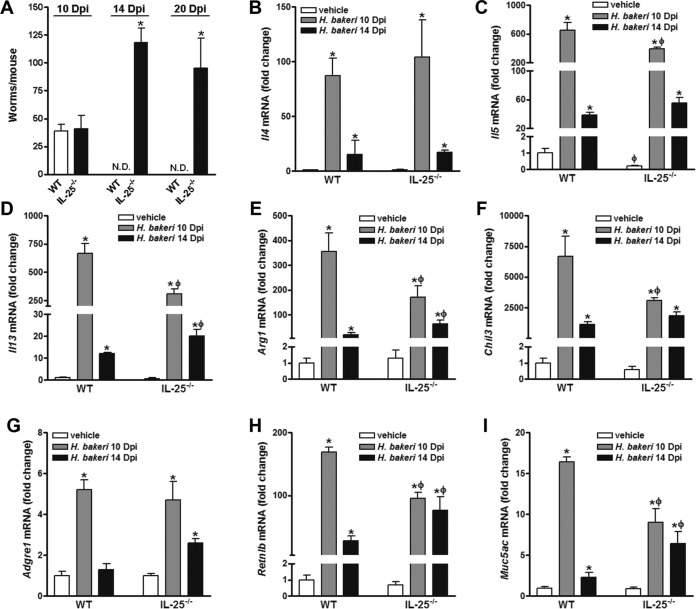
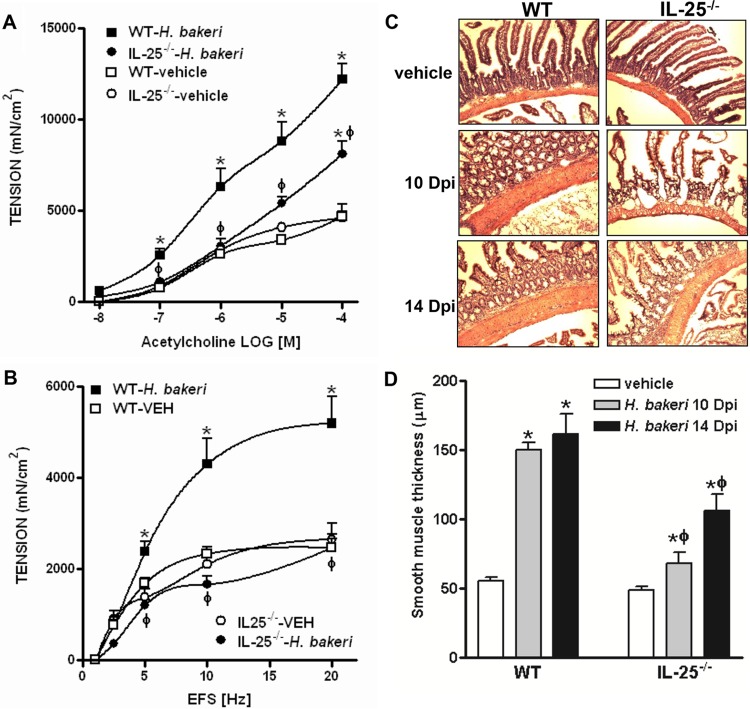
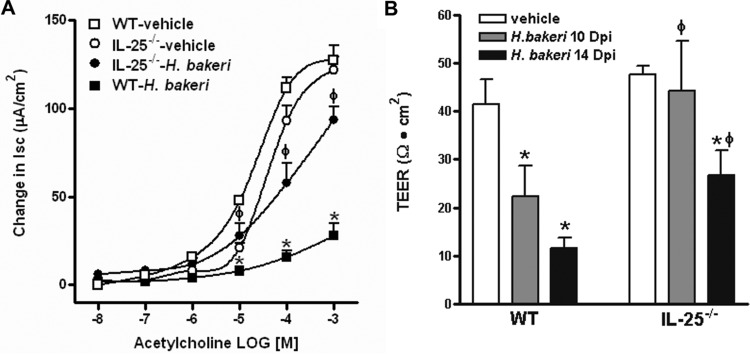
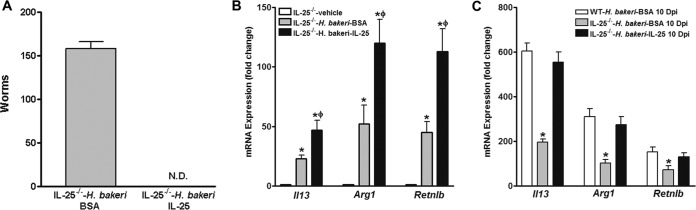
Infection with parasitic nematodes, especially gastrointestinal geohelminths, affects hundreds of millions of people worldwide and thus poses a major risk to global health. The host mechanism of defense against enteric nematode infection remains to be fully understood, but it involves a polarized type 2 immunity leading to alterations in intestinal function that facilitate worm expulsion. We investigated the role of interleukin-25 (IL-25) in host protection against Heligmosomoides polygyrus bakeri infection in mice. Our results showed that Il25 and its receptor subunit, Il17rb, were upregulated during a primary infection and a secondary challenge infection with H. polygyrus bakeri Genetic deletion of IL-25 (IL-25-/-) led to an attenuated type 2 cytokine response and increased worm fecundity in mice with a primary H. polygyrus bakeri infection. In addition, the full spectrum of the host memory response against a secondary infection with H. polygyrus bakeri was severely impaired in IL-25-/- mice, including delayed type 2 cytokine responses, an attenuated functional response of the intestinal smooth muscle and epithelium, diminished intestinal smooth muscle hypertrophy/hyperplasia, and impaired worm expulsion. Furthermore, exogenous administration of IL-25 restored the host protective memory response against H. polygyrus bakeri infection in IL-25-/- mice. These data demonstrate that IL-25 is critical for host protective immunity against H. polygyrus bakeri infection, highlighting its potential application as a therapeutic agent against parasitic nematode infection worldwide.
Copyright © 2016, American Society for Microbiology. All Rights Reserved.
Figures
References
-
- Gerbe F, Sidot E, Smyth DJ, Ohmoto M, Matsumoto I, Dardalhon V, Cesses P, Garnier L, Pouzolles M, Brulin B, Bruschi M, Harcus Y, Zimmermann VS, Taylor N, Maizels RM, Jay P. 2016. Intestinal epithelial tuft cells initiate type 2 mucosal immunity to helminth parasites. Nature 529:226–230. doi:10.1038/nature16527. - DOI - PMC - PubMed
-
- Saenz SA, Siracusa MC, Monticelli LA, Ziegler CG, Kim BS, Brestoff JR, Peterson LW, Wherry EJ, Goldrath AW, Bhandoola A, Artis D. 2013. IL-25 simultaneously elicits distinct populations of innate lymphoid cells and multipotent progenitor type 2 (MPPtype2) cells. J Exp Med 210:1823–1837. doi:10.1084/jem.20122332. - DOI - PMC - PubMed
-
- Fallon PG, Ballantyne SJ, Mangan NE, Barlow JL, Dasvarma A, Hewett DR, McIlgorm A, Jolin HE, McKenzie AN. 2006. Identification of an interleukin (IL)-25-dependent cell population that provides IL-4, IL-5, and IL-13 at the onset of helminth expulsion. J Exp Med 203:1105–1116. doi:10.1084/jem.20051615. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
