

Insulin receptor substrate-1 time-dependently regulates bone formation by controlling collagen Iα2 expression via miR-342
- PMID: 27623927
- PMCID: PMC5102111
- DOI: 10.1096/fj.201600445RR
Insulin receptor substrate-1 time-dependently regulates bone formation by controlling collagen Iα2 expression via miR-342
Abstract
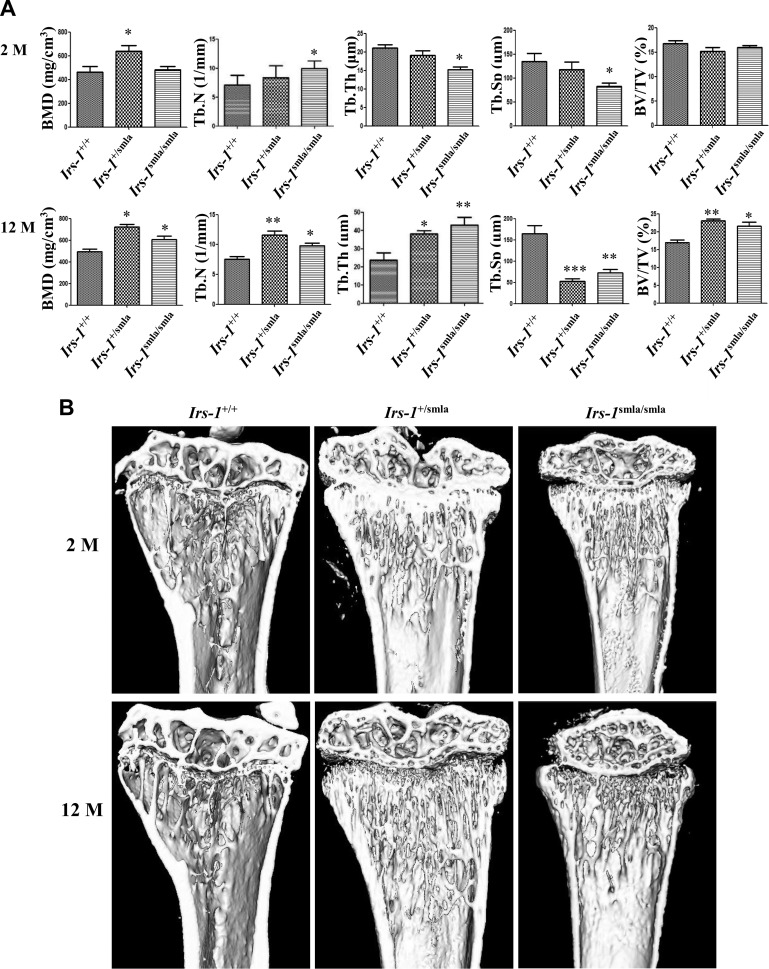
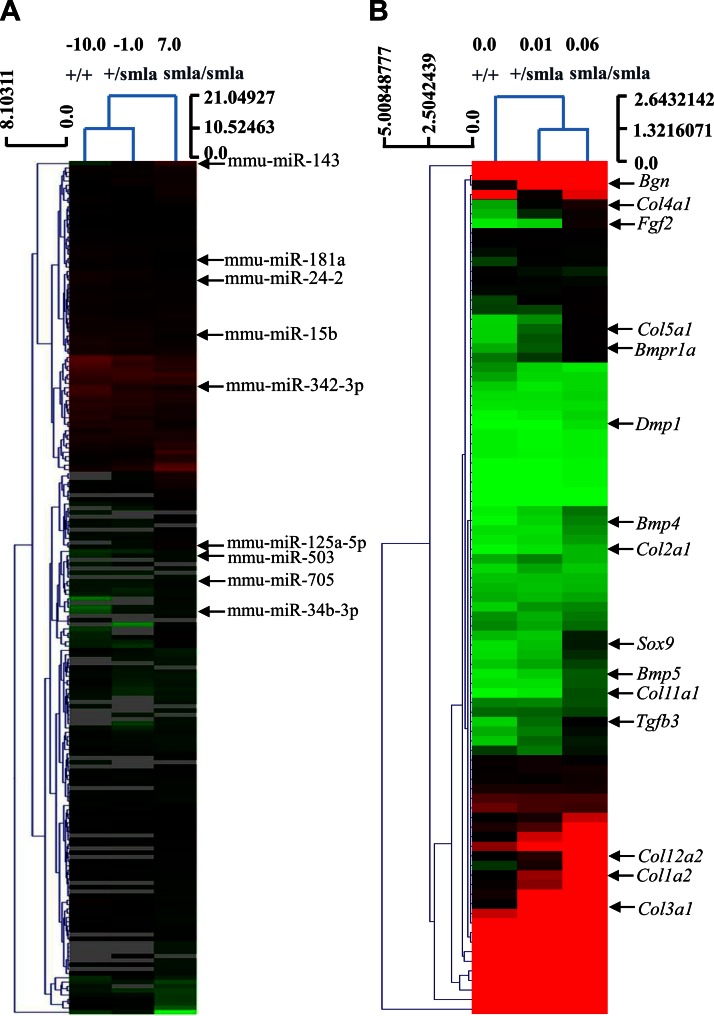
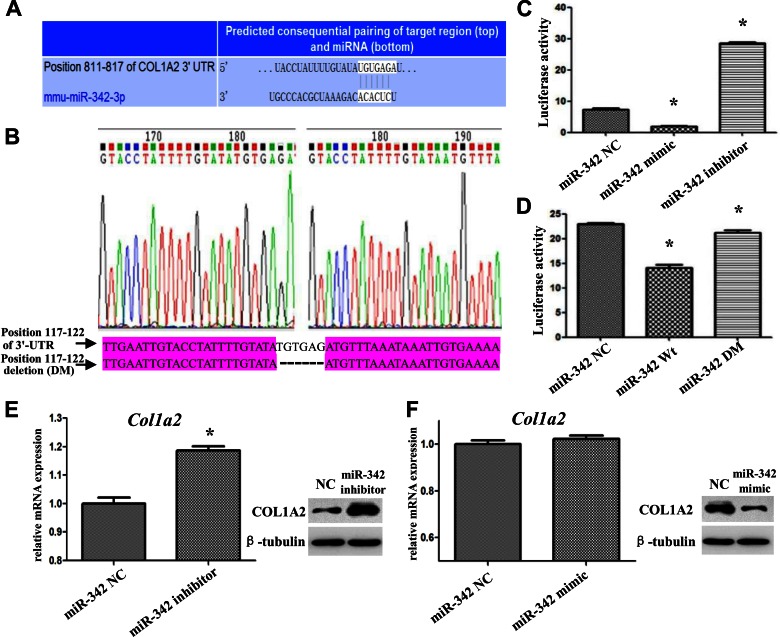
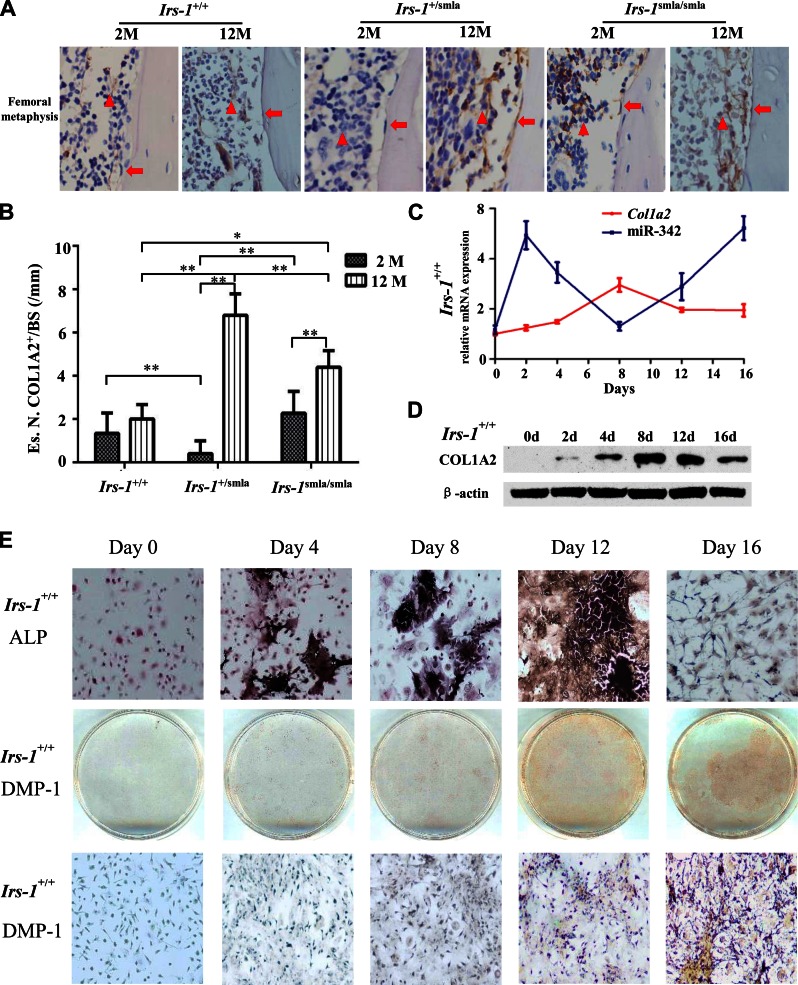
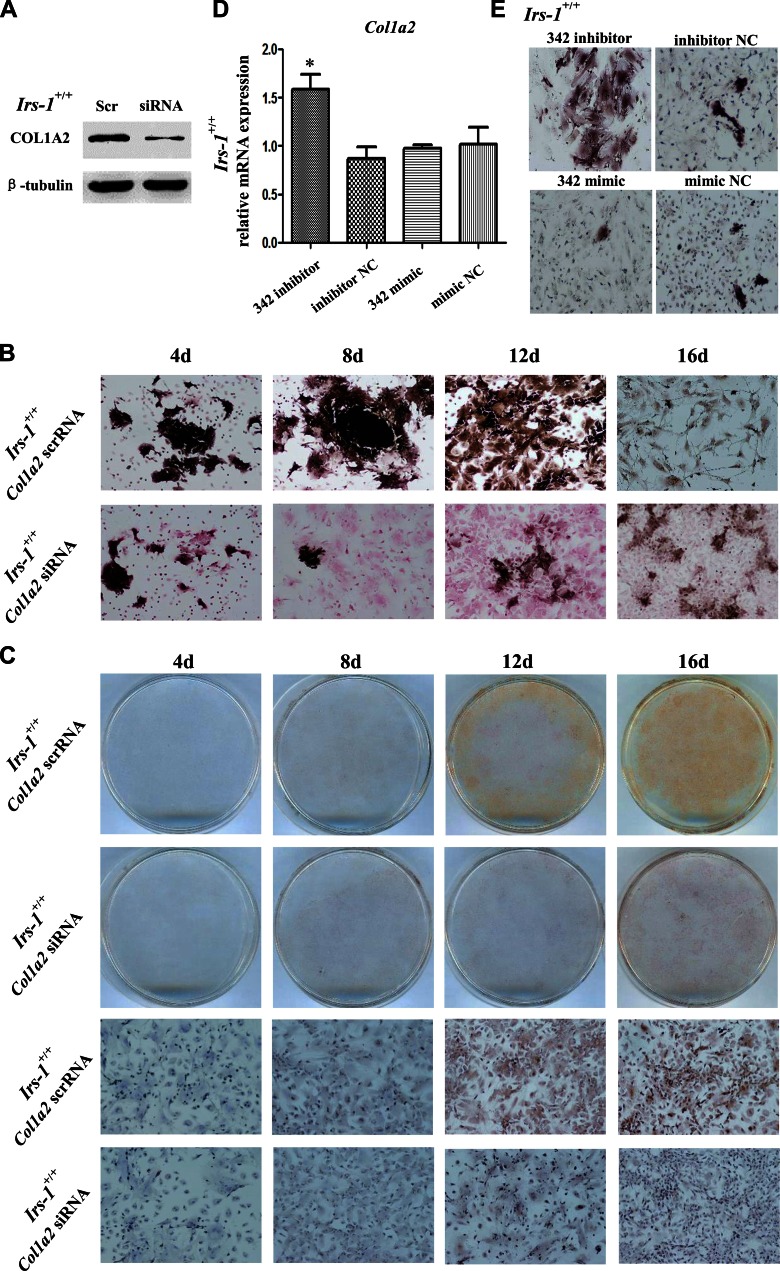
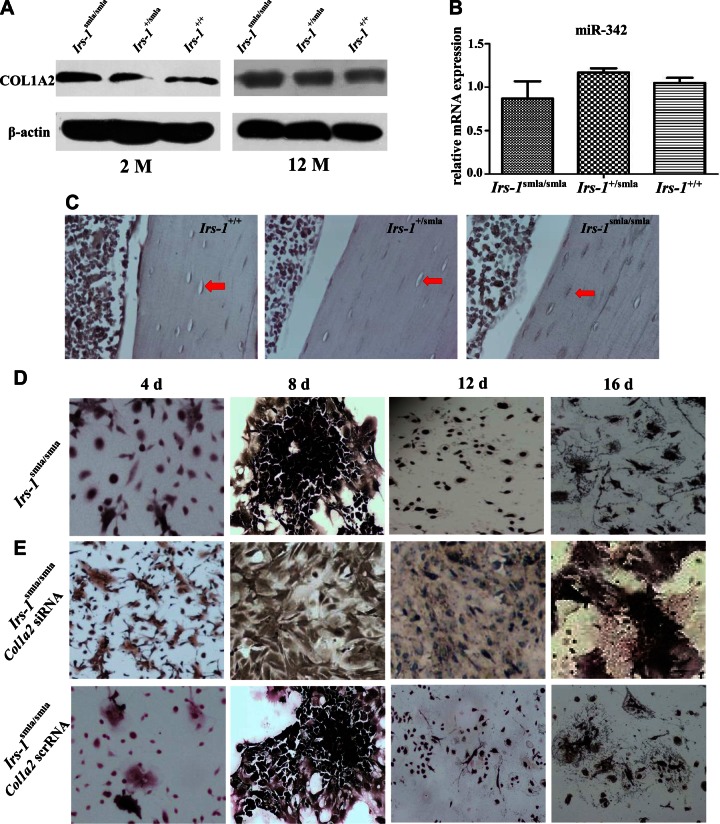
Insulin promotes bone formation via a well-studied canonical signaling pathway. An adapter in this pathway, insulin-receptor substrate (IRS)-1, has been implicated in the diabetic osteopathy provoked by impaired insulin signaling. To further investigate IRS-1's role in the bone metabolism, we generated Irs-1-deficient Irs-1smla/smla mice. These null mice developed a spontaneous mutation that led to an increase in trabecular thickness (Tb.Th) in 12-mo-old, but not in 2-mo-old mice. Analyses of the bone marrow stromal cells (BMSCs) from these mice revealed their differential expression of osteogenesis-related genes and miRNAs. The expression of miR-342, predicted and then proven to target the gene encoding collagen type Iα2 (COL1A2), was reduced in BMSCs derived from Irs-1-null mice. COL1A2 expression was then shown to be age dependent in osteoblasts and BMSCs derived from Irs-1smla/smla mice. After the induction of osteogenesis in BMSCs, miR-342 expression correlated inversely with that of Col1a2 Further, Col1a2-specific small interfering RNA (siRNA) reduced alkaline phosphatase (ALP) activity and inhibited BMSC differentiation into osteocyte-like cells, both in wild-type (WT) and Irs-1smla/smla mice. Conversely, in Irs-1smla/smla osteocytes overexpressing COL1A2, ALP-positive staining was stronger than in WT osteocytes. In summary, we uncovered a temporal regulation of BMSC differentiation/bone formation, controlled via Irs-1/miR-342 mediated regulation of Col1a2 expression.-Guo, Y., Tang, C.-Y., Man, X.-F., Tang, H.-N., Tang, J., Wang, F., Zhou, C.-L., Tan, S.-W., Feng, Y.-Z., Zhou, H.-D. Insulin receptor substrate-1 time-dependently regulates bone formation by controlling collagen Iα2 expression via miR-342.
Keywords: bone formation; collagen 1α2; insulin receptor substrate; miR-342.
© The Author(s).
Figures
References
-
- Dhaliwal R., Cibula D., Ghosh C., Weinstock R. S., Moses A. M. (2014) Bone quality assessment in type 2 diabetes mellitus. Osteoporos. Int. 25, 1969–1973 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
