

Development of a reliable automated screening system to identify small molecules and biologics that promote human β-cell regeneration
- PMID: 27624103
- PMCID: PMC5130356
- DOI: 10.1152/ajpendo.00515.2015
Development of a reliable automated screening system to identify small molecules and biologics that promote human β-cell regeneration
Abstract
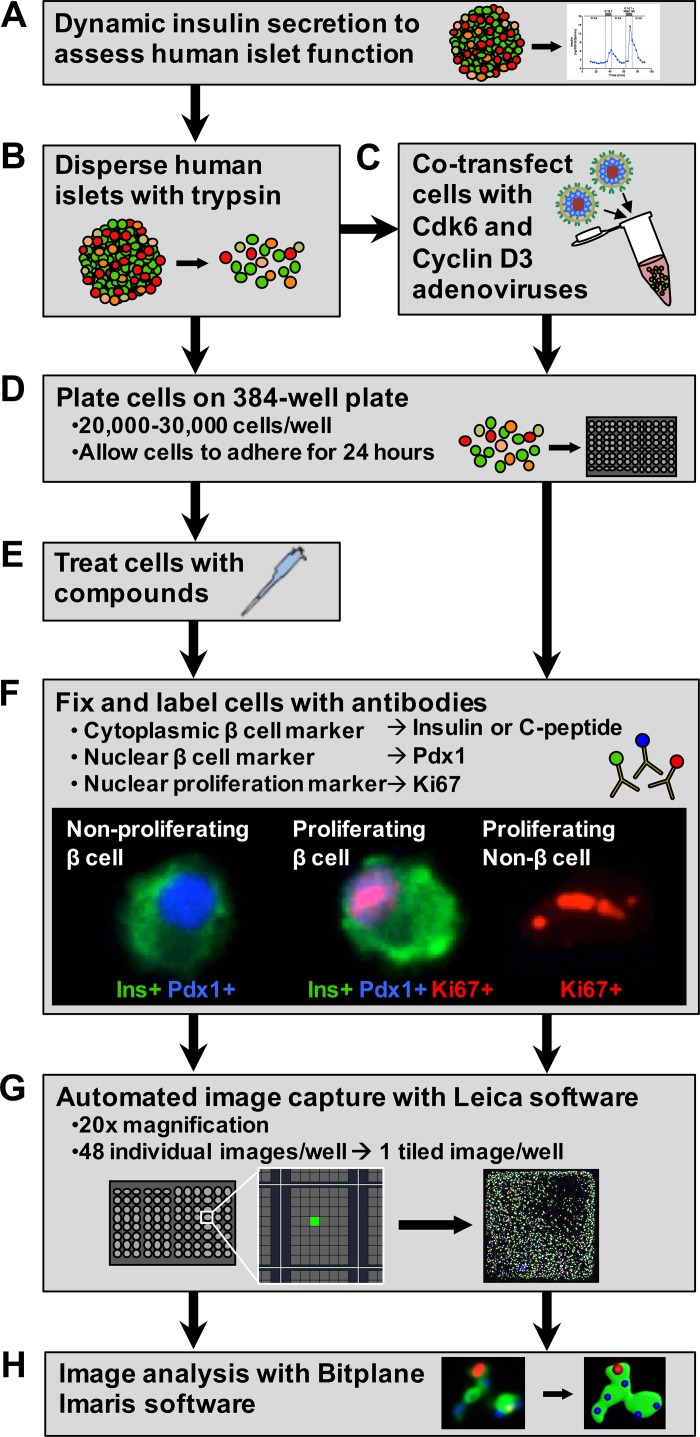
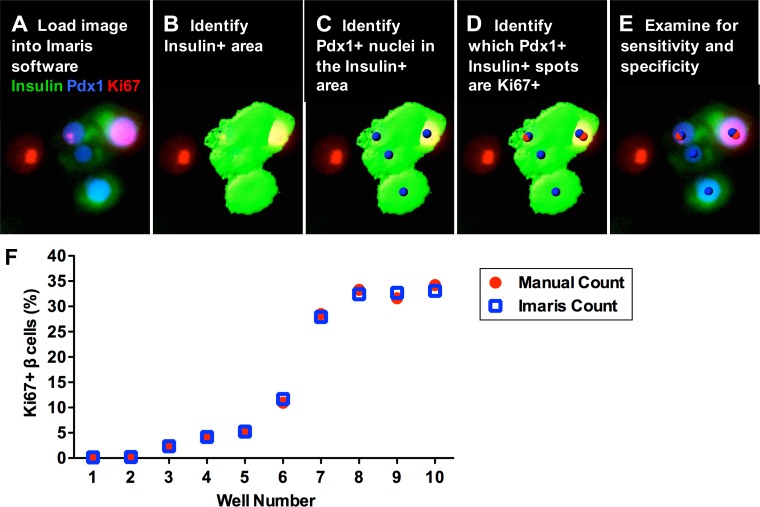
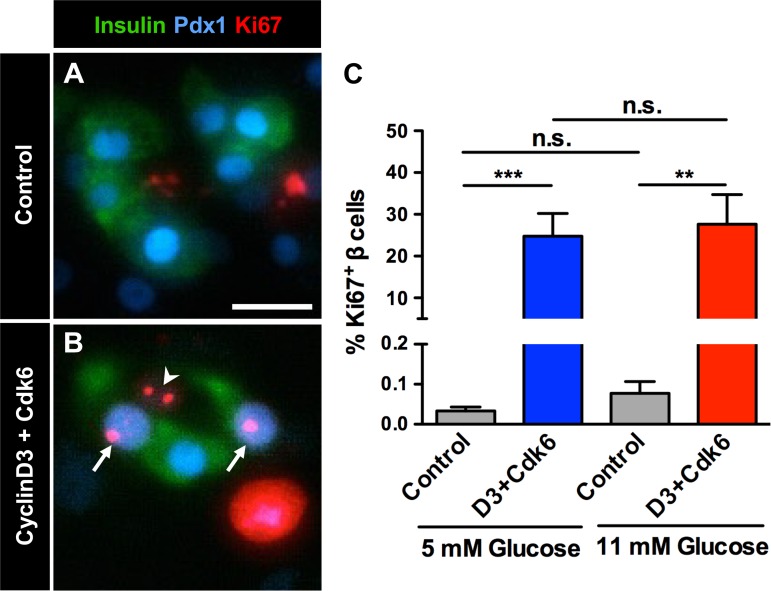
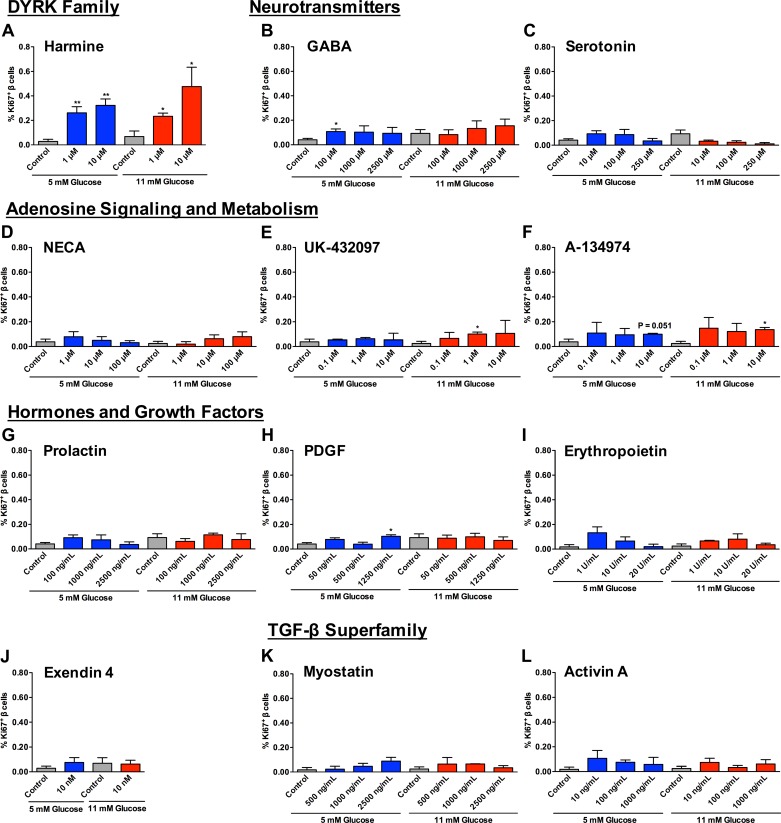
Numerous compounds stimulate rodent β-cell proliferation; however, translating these findings to human β-cells remains a challenge. To examine human β-cell proliferation in response to such compounds, we developed a medium-throughput in vitro method of quantifying adult human β-cell proliferation markers. This method is based on high-content imaging of dispersed islet cells seeded in 384-well plates and automated cell counting that identifies fluorescently labeled β-cells with high specificity using both nuclear and cytoplasmic markers. β-Cells from each donor were assessed for their function and ability to enter the cell cycle by cotransduction with adenoviruses encoding cell cycle regulators cdk6 and cyclin D3. Using this approach, we tested 12 previously identified mitogens, including neurotransmitters, hormones, growth factors, and molecules, involved in adenosine and Tgf-1β signaling. Each compound was tested in a wide concentration range either in the presence of basal (5 mM) or high (11 mM) glucose. Treatment with the control compound harmine, a Dyrk1a inhibitor, led to a significant increase in Ki-67+ β-cells, whereas treatment with other compounds had limited to no effect on human β-cell proliferation. This new scalable approach reduces the time and effort required for sensitive and specific evaluation of human β-cell proliferation, thus allowing for increased testing of candidate human β-cell mitogens.
Keywords: human islet; β-cell proliferation.
Figures
References
-
- Beattie GM, Cirulli V, Lopez AD, Hayek A. Ex vivo expansion of human pancreatic endocrine cells. J Clin Endocrinol Metab 82: 1852–1856, 1997. - PubMed
-
- Brelje TC, Stout LE, Bhagroo NV, Sorenson RL. Distinctive roles for prolactin and growth hormone in the activation of signal transducer and activator of transcription 5 in pancreatic islets of langerhans. Endocrinology 145: 4162–4175, 2004. - PubMed
-
- Brissova M, Fowler MJ, Nicholson WE, Chu A, Hirshberg B, Harlan DM, Powers AC. Assessment of human pancreatic islet architecture and composition by laser scanning confocal microscopy. J Histochem Cytochem 53: 1087–1097, 2005. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources