

Transcriptional Regulation of Brain-derived Neurotrophic Factor Coding Exon IX: ROLE OF NUCLEAR RESPIRATORY FACTOR 2
- PMID: 27624937
- PMCID: PMC5077195
- DOI: 10.1074/jbc.M116.742304
Transcriptional Regulation of Brain-derived Neurotrophic Factor Coding Exon IX: ROLE OF NUCLEAR RESPIRATORY FACTOR 2
Abstract
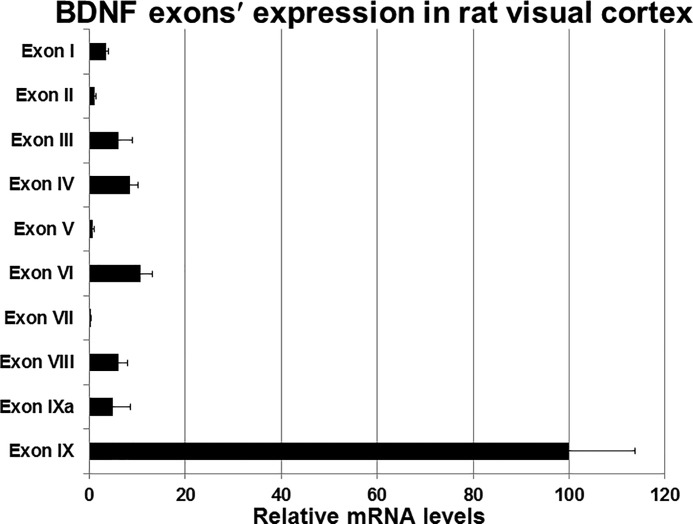
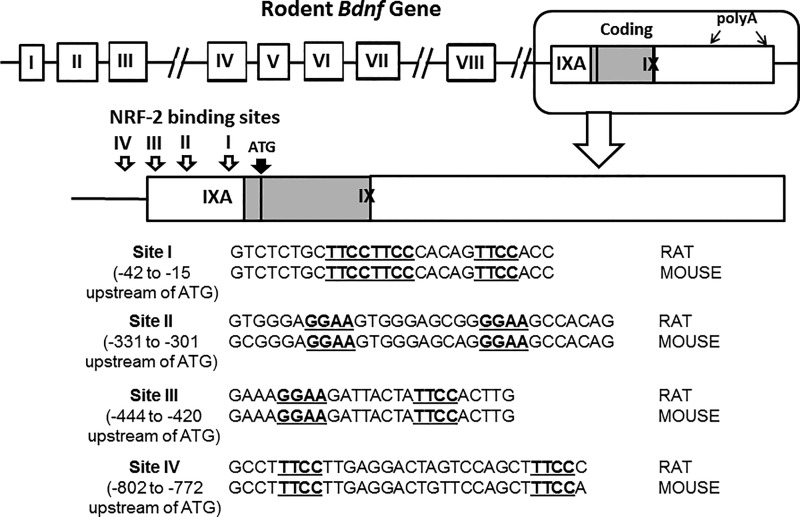
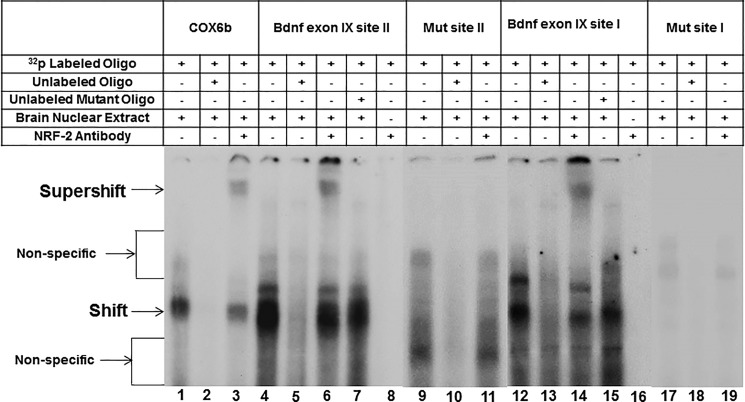
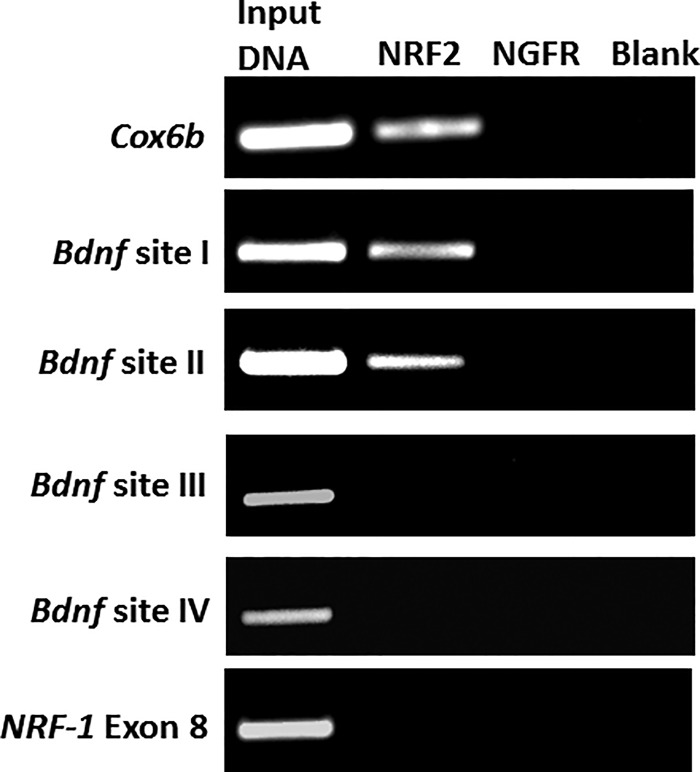
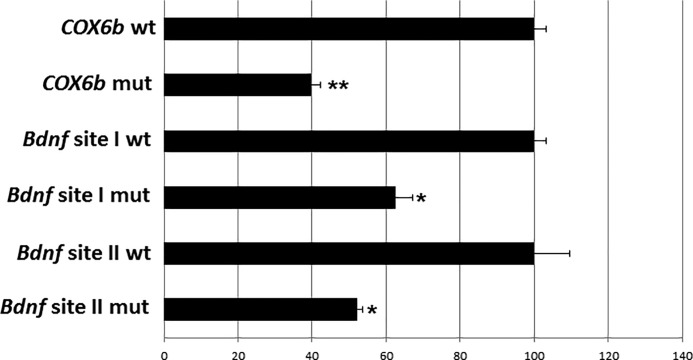
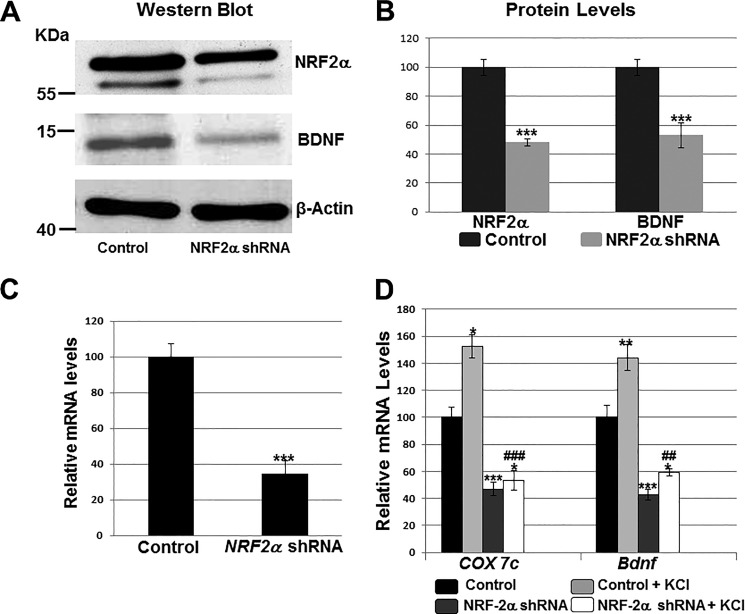
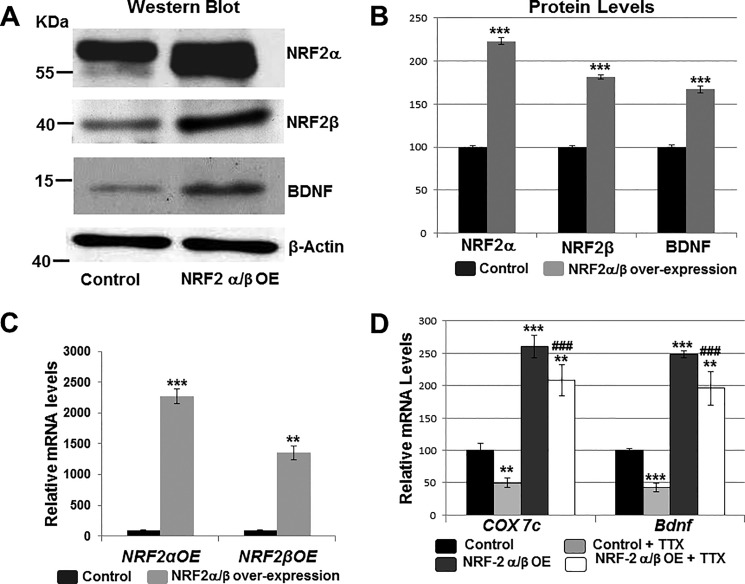
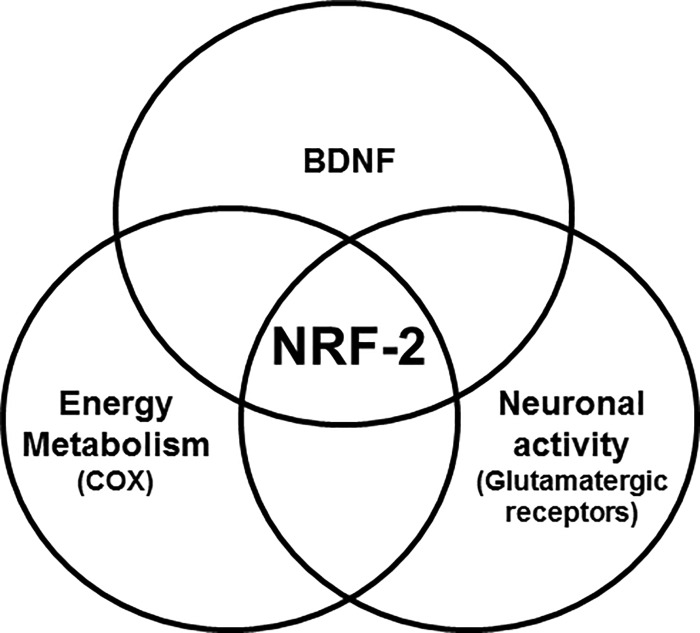
Brain-derived neurotrophic factor (BDNF) is an active neurotrophin abundantly expressed throughout the nervous system. It plays an important role in synaptic transmission, plasticity, neuronal proliferation, differentiation, survival, and death. The Bdnf gene in rodents has eight non-coding exons and only a single coding exon (IX). Despite its recognized regulation by neuronal activity, relatively little is known about its transcriptional regulation, and even less about the transcription factor candidates that may play such a role. The goal of the present study was to probe for such a candidate that may regulate exon IX in the rat Bdnf gene. Our in silico analysis revealed tandem binding sites for nuclear respiratory factor 2 (NRF-2) on the promoter of exon IX. NRF-2 is of special significance because it co-regulates the expressions of mediators of energy metabolism (cytochrome c oxidase) and mediators of neuronal activity (glutamatergic receptors). To test our hypothesis that NRF-2 also regulates the Bdnf gene, we performed electrophoretic mobility shift assay (EMSA), chromatin immunoprecipitation (ChIP), promoter cloning, and site-directed mutagenesis, real-time quantitative PCR (RT-qPCR), and Western blotting analysis. Results indicate that NRF-2 functionally regulates exon IX of the rat Bdnf gene. The binding sites of NRF-2 are conserved between rats and mice. Overexpressing NRF-2 up-regulated the expression of Bdnf exon IX, whereas knocking down NRF-2 down-regulated such expression. These findings are consistent with our hypothesis that NRF-2, in addition to regulating the coupling between neuronal activity and energy metabolism, also regulates the expression of BDNF, which is intimately associated with energy-demanding neuronal activity.
Keywords: brain-derived neurotrophic factor (BDNF); chromatin immunoprecipitation (ChiP); short hairpin RNA (shRNA); transcription factor; transcription regulation.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Ernfors P., Wetmore C., Olson L., and Persson H. (1990) Identification of cells in rat brain and peripheral tissues expressing mRNA for members of the nerve growth factor family. Neuron 5, 511–526 - PubMed
-
- Katoh-Semba R., Takeuchi I. K., Semba R., and Kato K. (1997) Distribution of brain-derived neurotrophic factor in rats and its changes with development in the brain. J. Neurochem. 69, 34–42 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
