

Regulation of Herpes Simplex Virus Glycoprotein-Induced Cascade of Events Governing Cell-Cell Fusion
- PMID: 27630245
- PMCID: PMC5110162
- DOI: 10.1128/JVI.01501-16
Regulation of Herpes Simplex Virus Glycoprotein-Induced Cascade of Events Governing Cell-Cell Fusion
Abstract
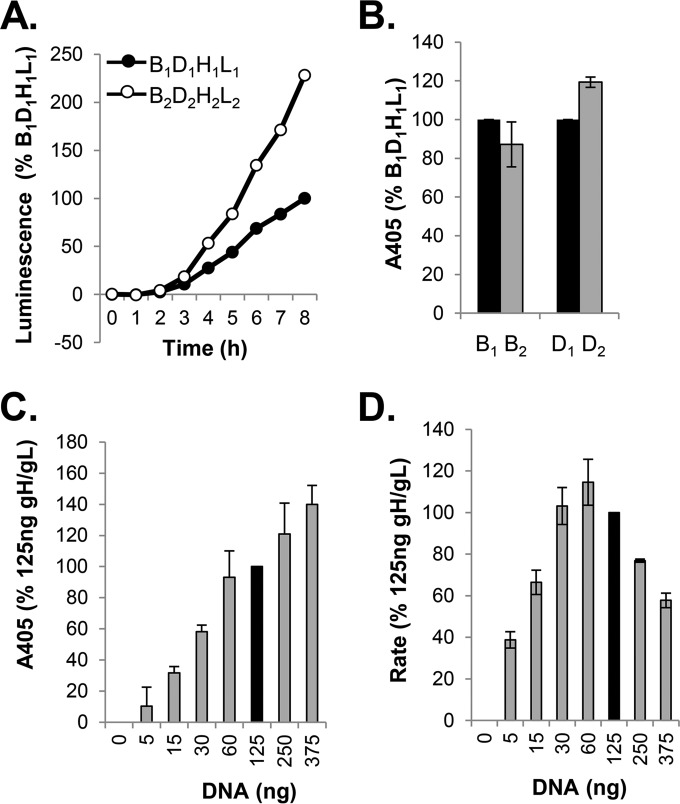
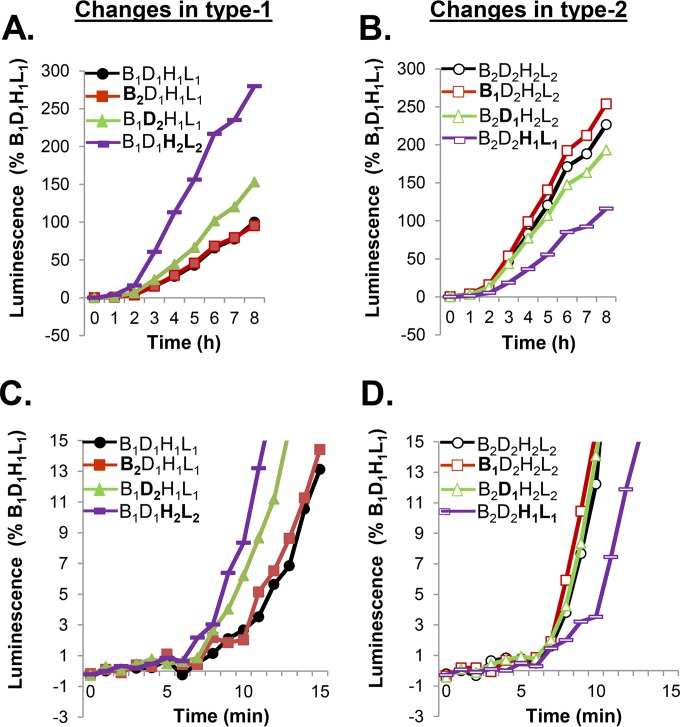
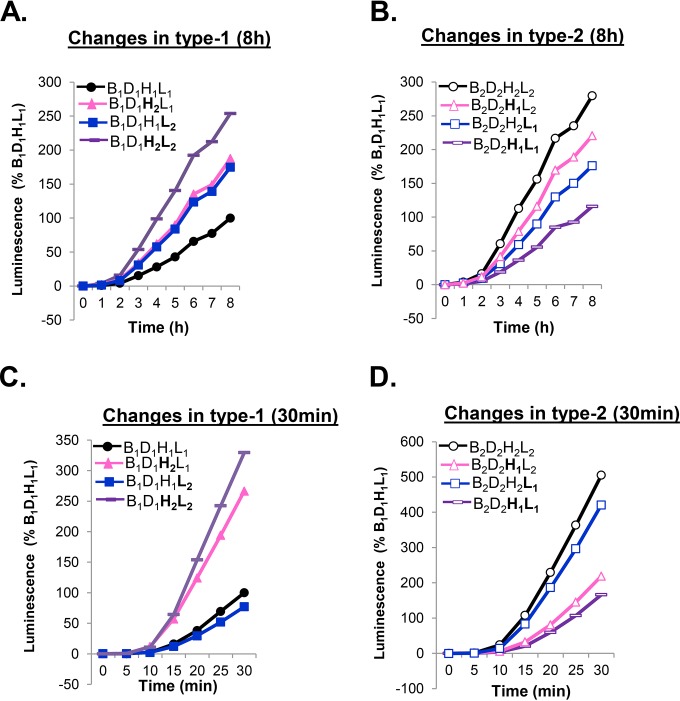
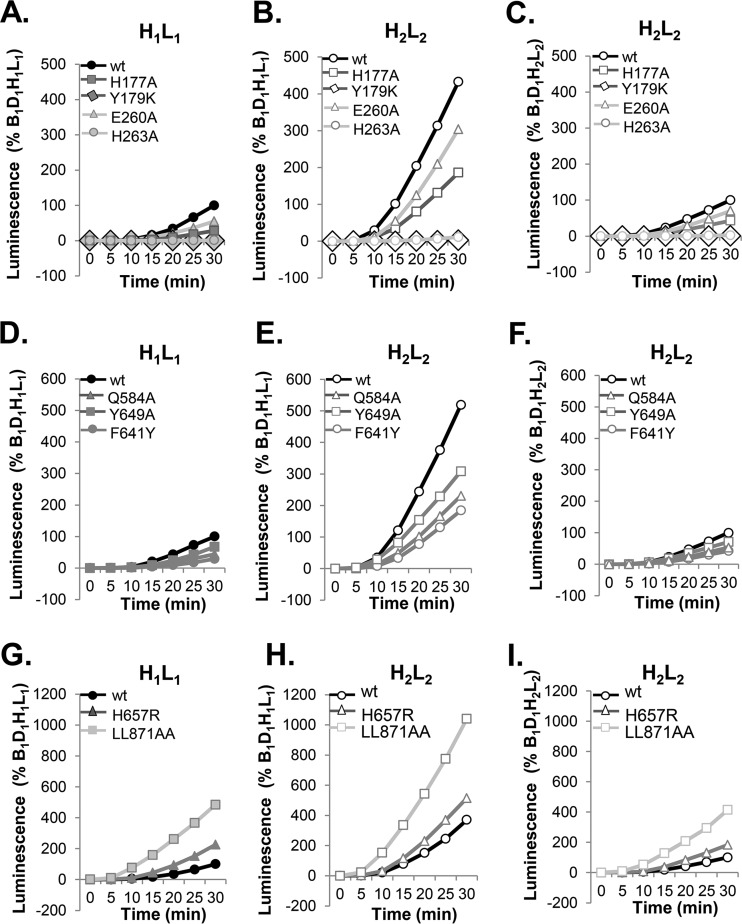
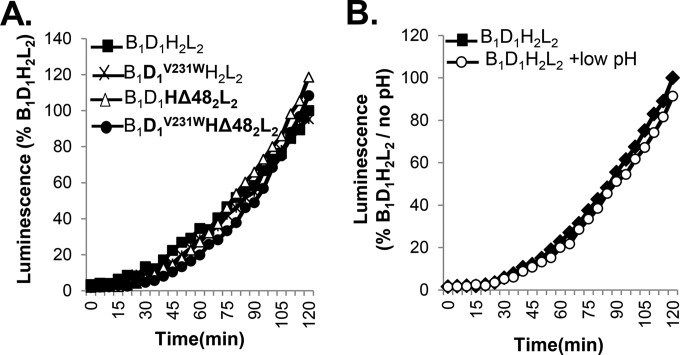
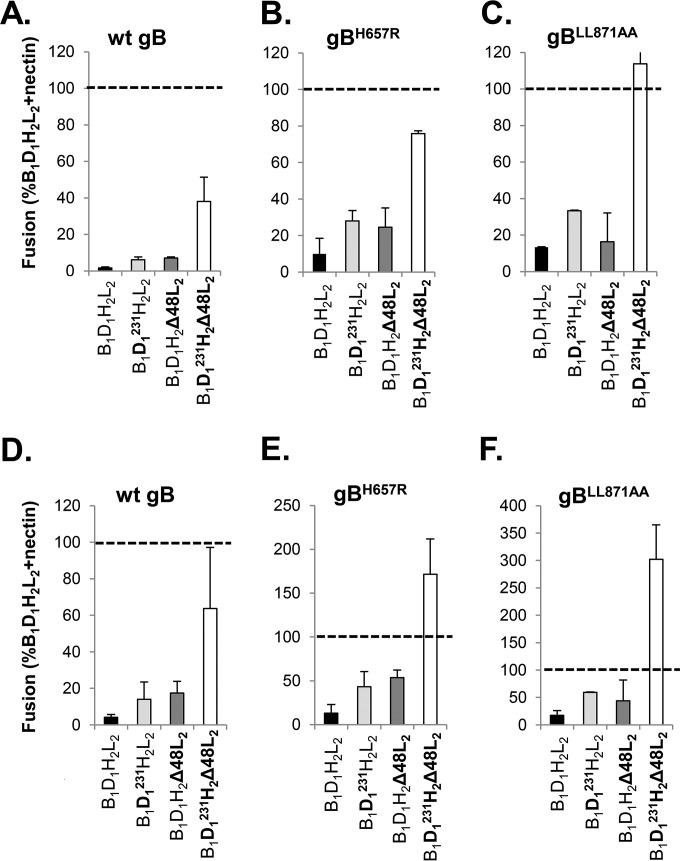
Receptor-dependent herpes simplex virus (HSV)-induced cell-cell fusion requires glycoproteins gD, gH/gL, and gB. Our current model posits that during fusion, receptor-activated conformational changes in gD activate gH/gL, which subsequently triggers the transformation of the prefusion form of gB into a fusogenic state. To examine the role of each glycoprotein in receptor-dependent cell-cell fusion, we took advantage of our discovery that fusion by wild-type herpes simplex virus 2 (HSV-2) glycoproteins occurs twice as fast as that achieved by HSV-1 glycoproteins. By sequentially swapping each glycoprotein between the two serotypes, we established that fusion speed was governed by gH/gL, with gH being the main contributor. While the mutant forms of gB fuse at distinct rates that are dictated by their molecular structure, these restrictions can be overcome by gH/gL of HSV-2 (gH2/gL2), thereby enhancing their activity. We also found that deregulated forms of gD of HSV-1 (gD1) and gH2/gL2 can alter the fusogenic potential of gB, promoting cell fusion in the absence of a cellular receptor, and that deregulated forms of gB can drive the fusion machinery to even higher levels. Low pH enhanced fusion by affecting the structure of both gB and gH/gL mutants. Together, our data highlight the complexity of the fusion machinery, the impact of the activation state of each glycoprotein on the fusion process, and the critical role of gH/gL in regulating HSV-induced fusion. IMPORTANCE Cell-cell fusion mediated by HSV glycoproteins requires gD, gH/gL, gB, and a gD receptor. Here, we show that fusion by wild-type HSV-2 glycoproteins occurs twice as fast as that achieved by HSV-1 glycoproteins. By sequentially swapping each glycoprotein between the two serotypes, we found that the fusion process was controlled by gH/gL. Restrictions imposed on the gB structure by mutations could be overcome by gH2/gL2, enhancing the activity of the mutants. Under low-pH conditions or when using deregulated forms of gD1 and gH2/gL2, the fusogenic potential of gB could only be increased in the absence of receptor, underlining the exquisite regulation that occurs in the presence of receptor. Our data highlight the complexity of the fusion machinery, the impact of the activation state of each glycoprotein on the fusion process, and the critical role of gH/gL in regulating HSV-induced fusion.
Copyright © 2016, American Society for Microbiology. All Rights Reserved.
Figures
References
-
- Baines JD, Pellett PE. 2007. Genetic comparison of human alphaherpesvirus genomes, p 61–69. In Arvin A, Campadelli-Fiume G, Mocarski E, Moore PS, Roizman B, Whitley R, Yamanishi K (ed), Human herpesviruses: biology, therapy, and immunoprophylaxis. Cambridge University Press, Cambridge, United Kingdom. - PubMed
-
- Roizman B, Taddeo B. 2007. The strategy of herpes simplex virus replication and takeover of the host cell, p 163–173. In Arvin A, Campadelli-Fiume G, Mocarski E, Moore PS, Roizman B, Whitley R, Yamanishi K (ed), Human herpesviruses: biology, therapy, and immunoprophylaxis. Cambridge University Press, Cambridge, United Kingdom. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
