

Accumulation of K+ in the synaptic cleft modulates activity by influencing both vestibular hair cell and calyx afferent in the turtle
- PMID: 27633787
- PMCID: PMC5285615
- DOI: 10.1113/JP273060
Accumulation of K+ in the synaptic cleft modulates activity by influencing both vestibular hair cell and calyx afferent in the turtle
Abstract
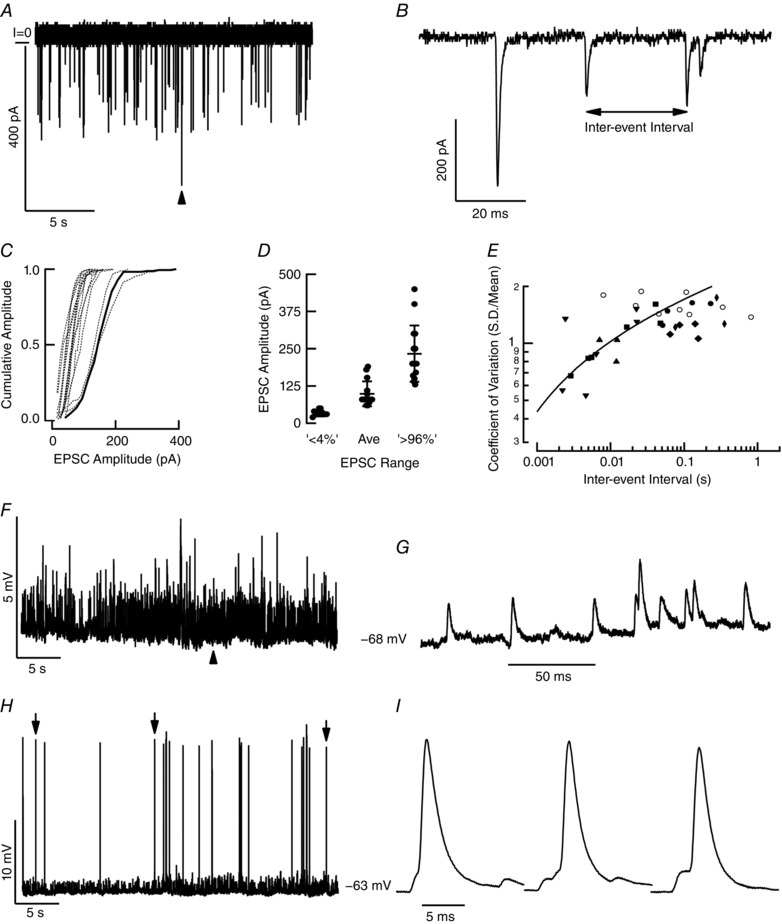
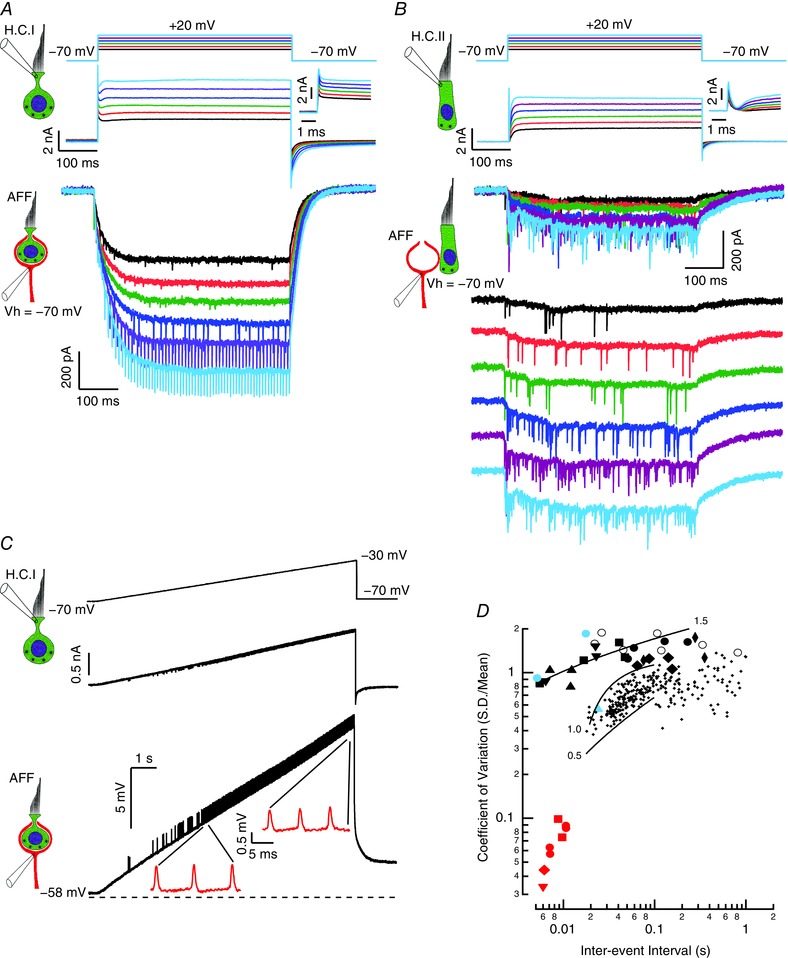
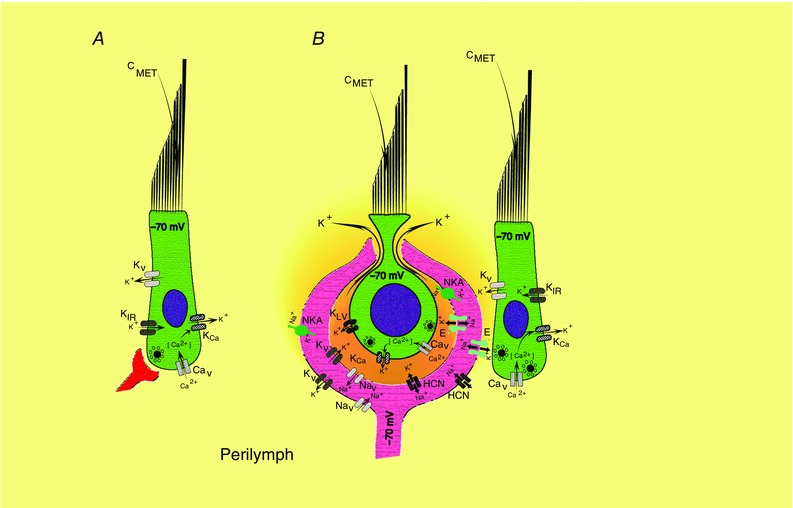
Key points: In the synaptic cleft between type I hair cells and calyceal afferents, K+ ions accumulate as a function of activity, dynamically altering the driving force and permeation through ion channels facing the synaptic cleft. High-fidelity synaptic transmission is possible due to large conductances that minimize hair cell and afferent time constants in the presence of significant membrane capacitance. Elevated potassium maintains hair cells near a potential where transduction currents are sufficient to depolarize them to voltages necessary for calcium influx and synaptic vesicle fusion. Elevated potassium depolarizes the postsynaptic afferent by altering ion permeation through hyperpolarization-activated cyclic nucleotide-gated (HCN) channels, and contributes to depolarizing the afferent to potentials where a single EPSP (quantum) can generate an action potential. With increased stimulation, hair cell depolarization increases the frequency of quanta released, elevates [K+ ]cleft and depolarizes the afferent to potentials at which smaller and smaller EPSPs would be sufficient to trigger APs.
Abstract: Fast neurotransmitters act in conjunction with slower modulatory effectors that accumulate in restricted synaptic spaces found at giant synapses such as the calyceal endings in the auditory and vestibular systems. Here, we used dual patch-clamp recordings from turtle vestibular hair cells and their afferent neurons to show that potassium ions accumulating in the synaptic cleft modulated membrane potentials and extended the range of information transfer. High-fidelity synaptic transmission was possible due to large conductances that minimized hair cell and afferent time constants in the presence of significant membrane capacitance. Increased potassium concentration in the cleft maintained the hair cell near potentials that promoted the influx of calcium necessary for synaptic vesicle fusion. The elevated potassium concentration also depolarized the postsynaptic neuron by altering ion permeation through hyperpolarization-activated cyclic nucleotide-gated (HCN) channels. This depolarization enabled the afferent to reliably generate action potentials evoked by single AMPA-dependent EPSPs. Depolarization of the postsynaptic afferent could also elevate potassium in the synaptic cleft, and would depolarize other hair cells enveloped by the same neuritic process increasing the fidelity of neurotransmission at those synapses as well. Collectively, these data demonstrate that neuronal activity gives rise to potassium accumulation, and suggest that potassium ion action on HCN channels can modulate neurotransmission, preserving the fidelity of high-speed synaptic transmission by dynamically shifting the resting potentials of both presynaptic and postsynaptic cells.
Keywords: excitatory synaptic transmission; hair cell; hyperpolarization-activated cyclic nucleotide-activated channels; potassium accumulation; synaptic modulation; vestibular system.
© 2016 The Authors. The Journal of Physiology © 2016 The Physiological Society.
Figures
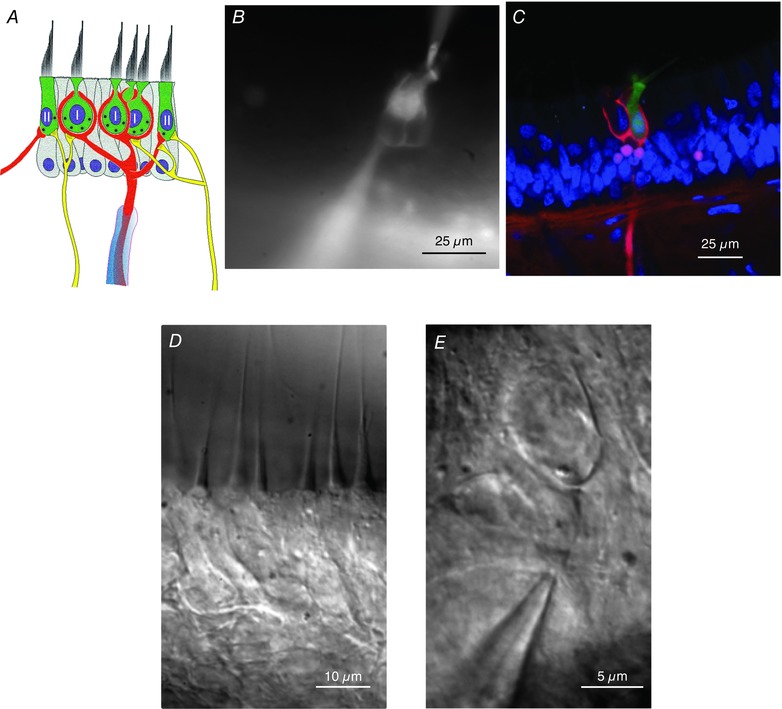
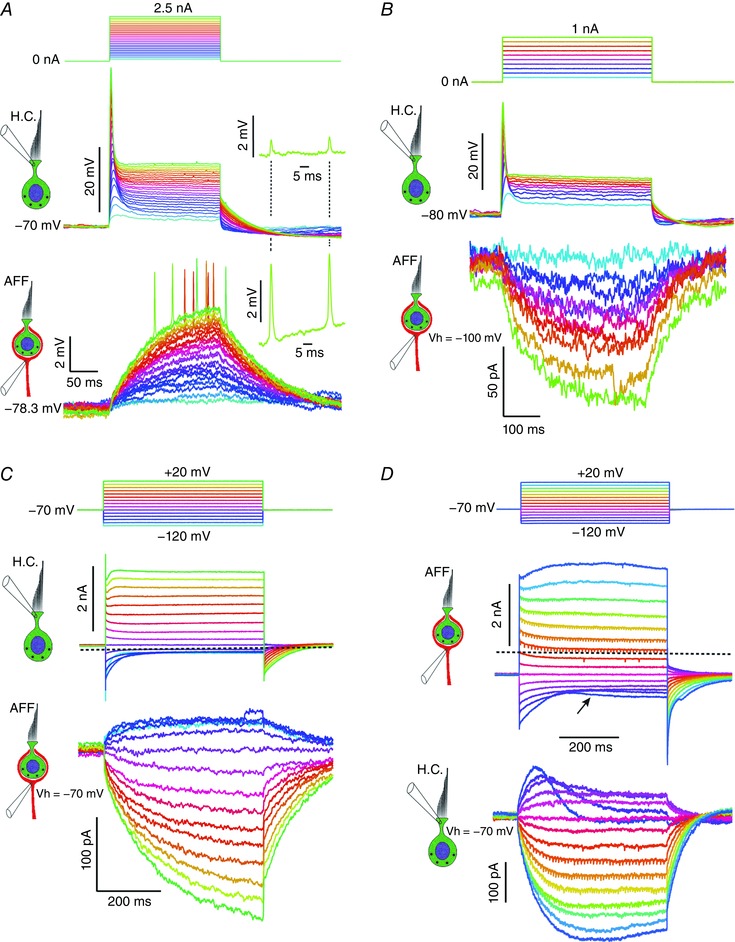
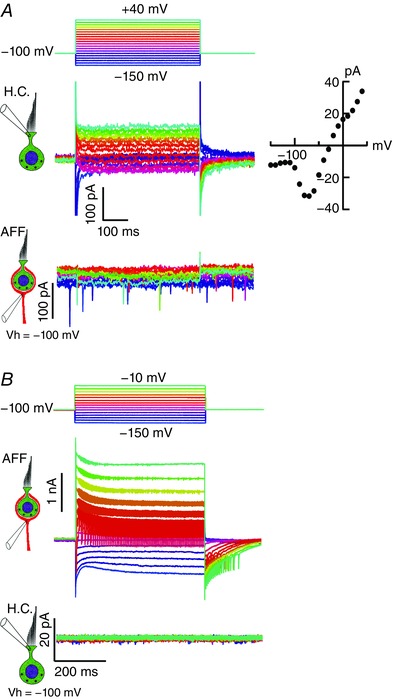
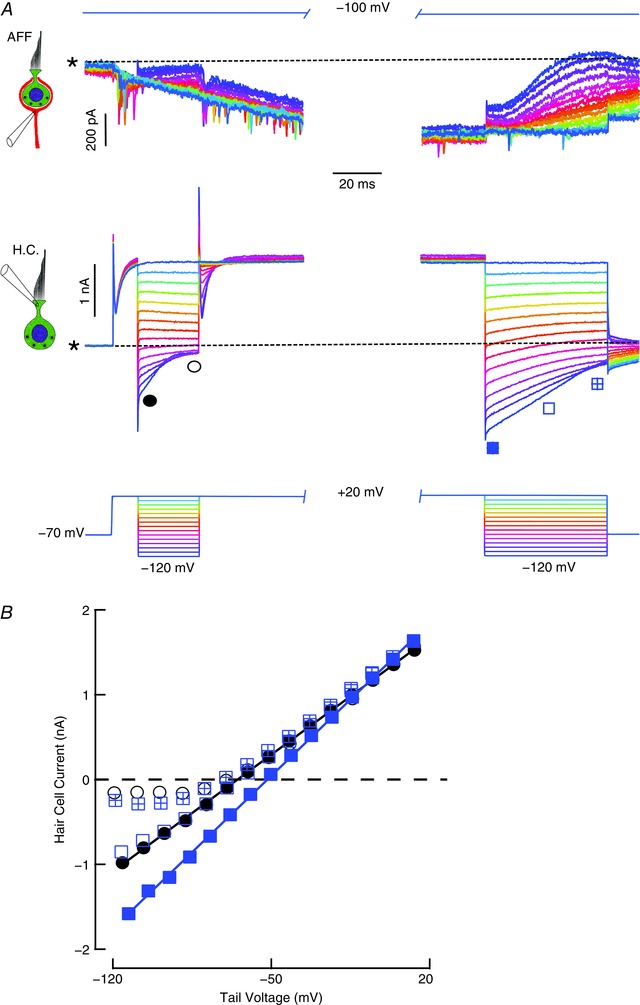
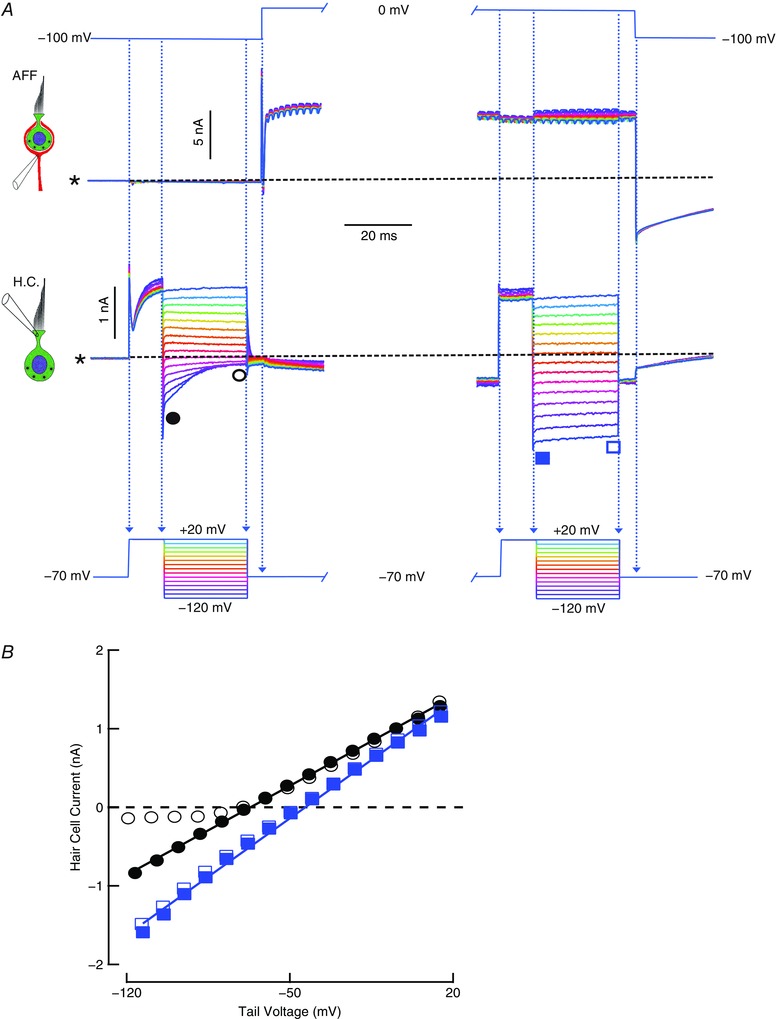
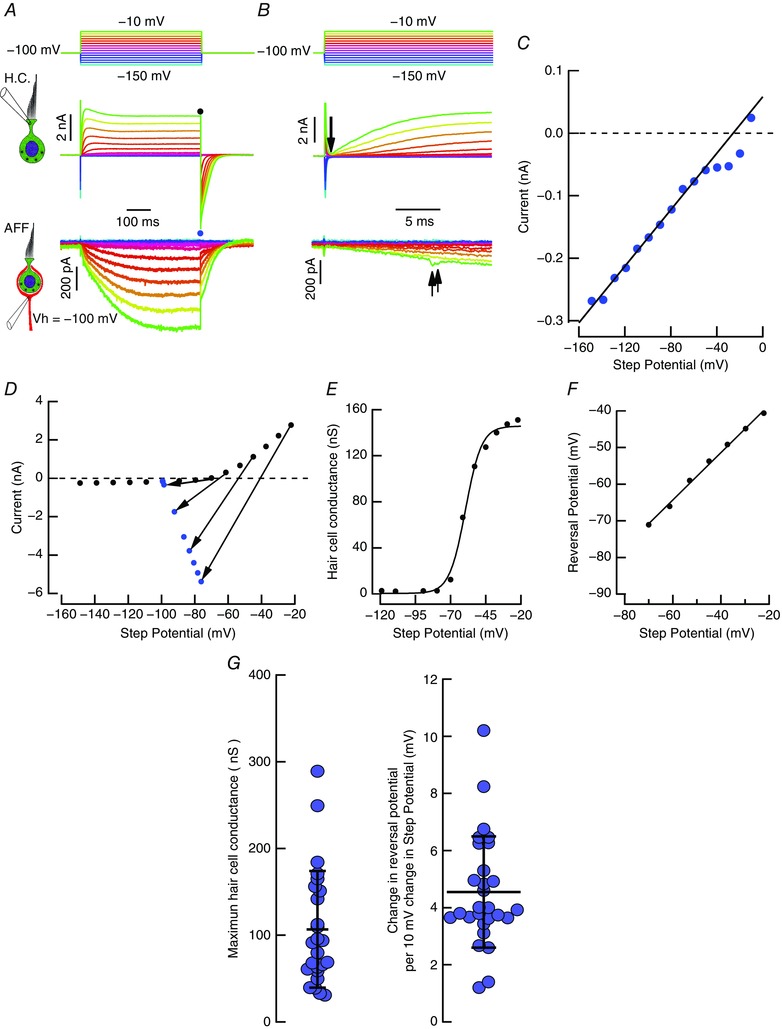
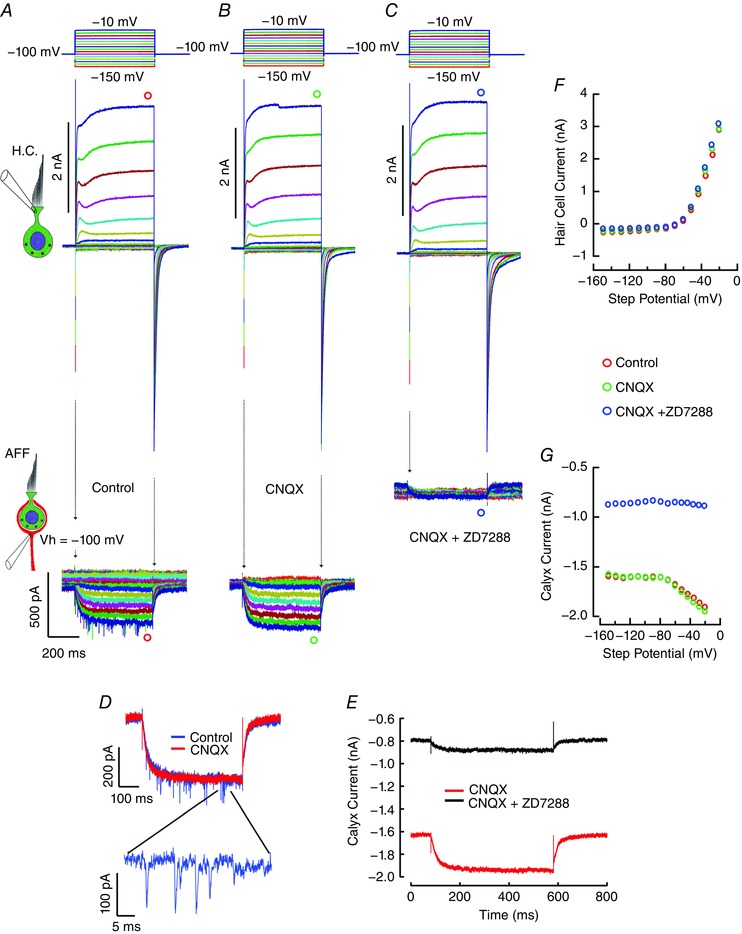
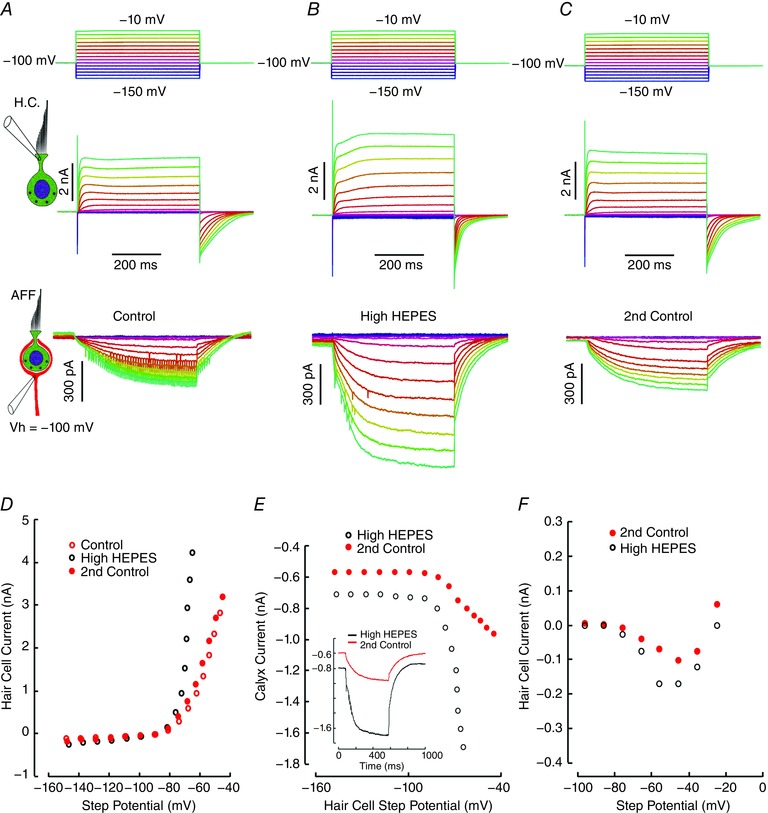
Comment in
-
Vestibular calyx, potassium: Kalium in Calyx Regnat.J Physiol. 2017 Feb 1;595(3):623. doi: 10.1113/JP273432. J Physiol. 2017. PMID: 28145016 Free PMC article. No abstract available.
References
-
- Adelman WJ Jr, Palti Y & Senft JP (1973). Potassium ion accumulation in a periaxonal space and its effect on the measurement of membrane potassium ion conductance. J Membr Biol 13, 387–410. - PubMed
-
- Araque A, Parpura V, Sanzgiri RP & Haydon PG (1999). Tripartite synapses: glia, the unacknowledged partner. Trends Neurosci 22, 208–215. - PubMed
-
- Attwell D, Cohen I & Eisner D (1979). Membrane potential and ion concentration stability conditions for a cell with a restricted extracellular space. Proc R Soc Lond B Biol Sci 206, 145–161. - PubMed
-
- Attwell D & Iles JF (1979). Synaptic transmission: ion concentration changes in the synaptic cleft. Proc R Soc Lond B Biol Sci 206, 115–131. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous