

Dopamine receptor DOP-4 modulates habituation to repetitive photoactivation of a C. elegans polymodal nociceptor
- PMID: 27634141
- PMCID: PMC5026203
- DOI: 10.1101/lm.041830.116
Dopamine receptor DOP-4 modulates habituation to repetitive photoactivation of a C. elegans polymodal nociceptor
Abstract
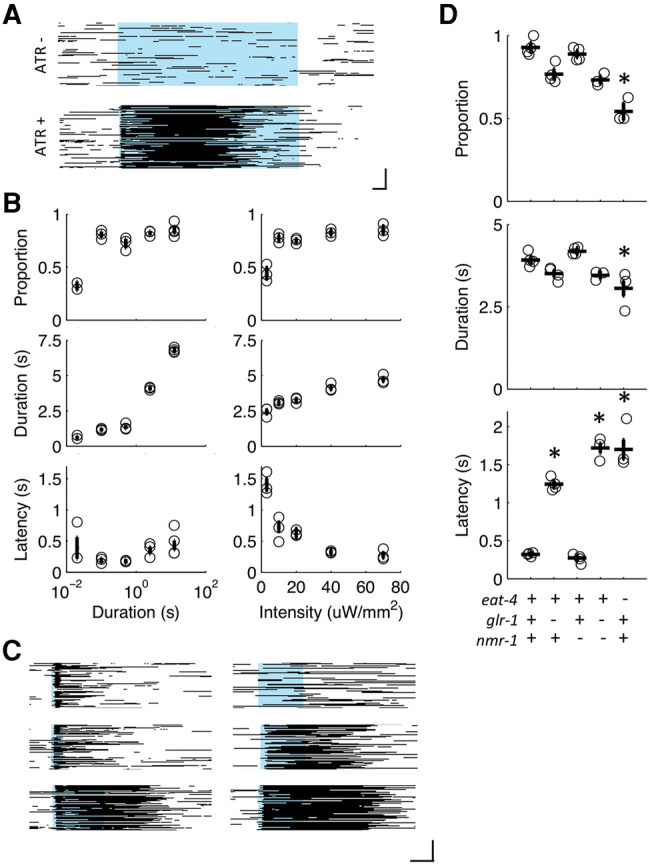
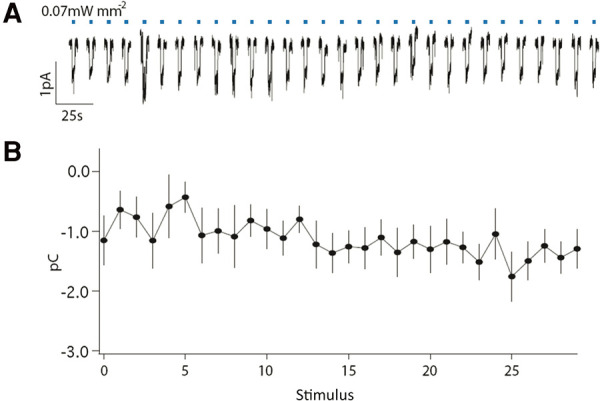
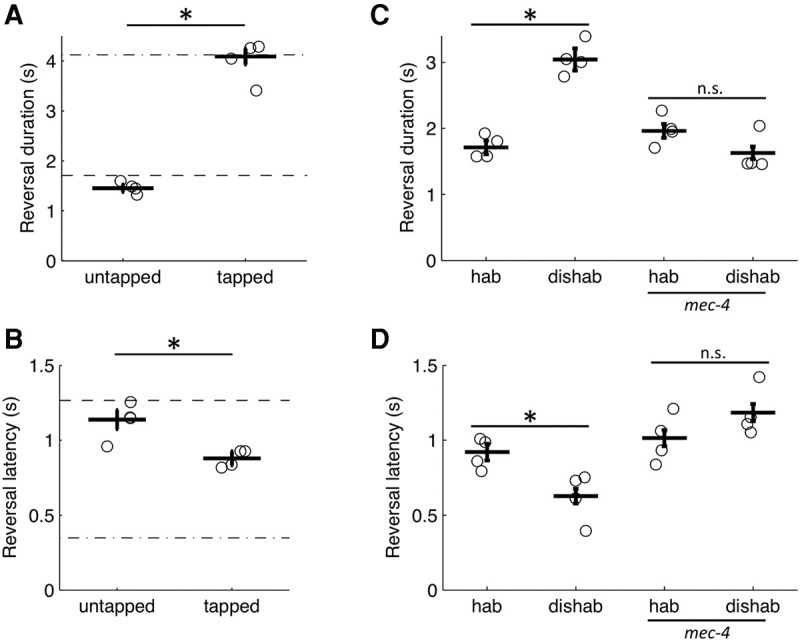
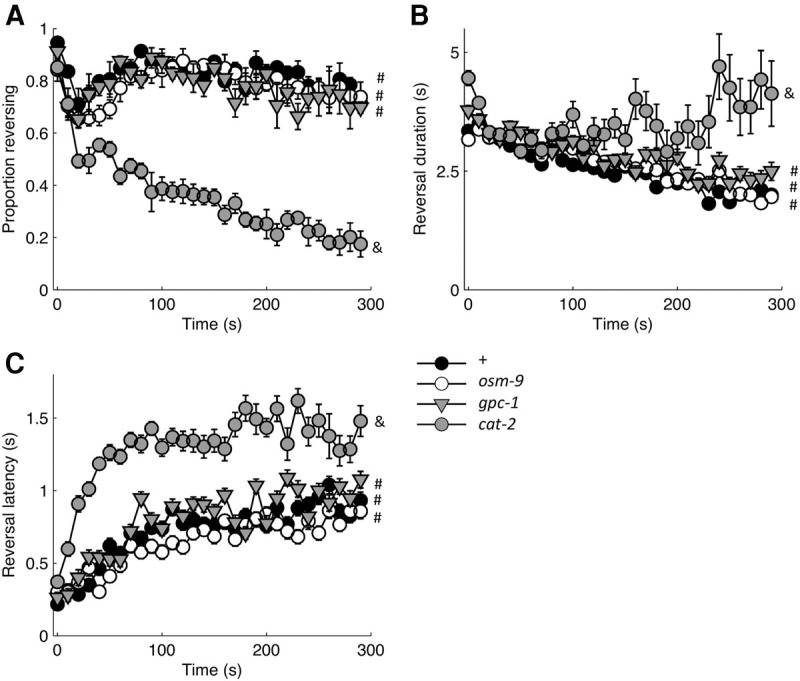
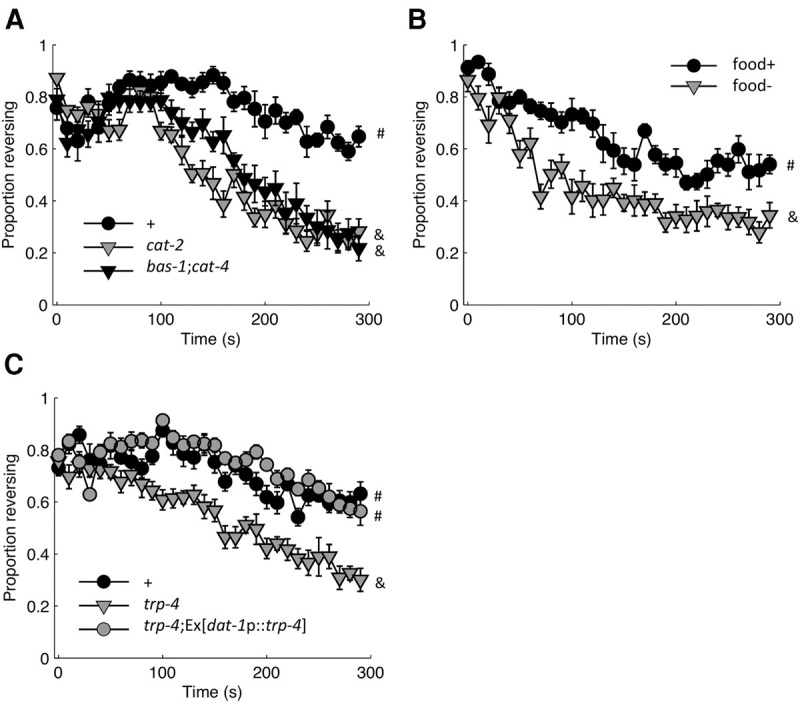
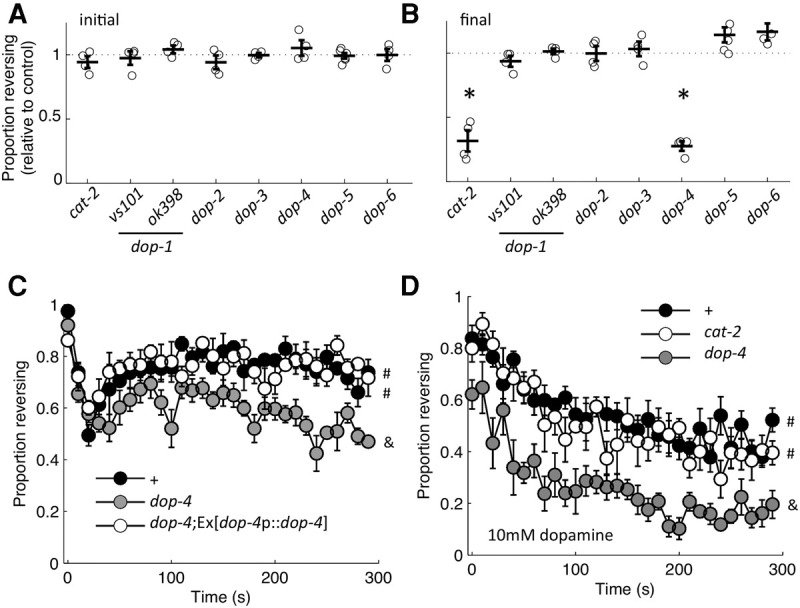
Habituation is a highly conserved phenomenon that remains poorly understood at the molecular level. Invertebrate model systems, like Caenorhabditis elegans, can be a powerful tool for investigating this fundamental process. Here we established a high-throughput learning assay that used real-time computer vision software for behavioral tracking and optogenetics for stimulation of the C. elegans polymodal nociceptor, ASH. Photoactivation of ASH with ChR2 elicited backward locomotion and repetitive stimulation altered aspects of the response in a manner consistent with habituation. Recording photocurrents in ASH, we observed no evidence for light adaptation of ChR2. Furthermore, we ruled out fatigue by demonstrating that sensory input from the touch cells could dishabituate the ASH avoidance circuit. Food and dopamine signaling slowed habituation downstream from ASH excitation via D1-like dopamine receptor, DOP-4. This assay allows for large-scale genetic and drug screens investigating mechanisms of nociception modulation.
© 2016 Ardiel et al.; Published by Cold Spring Harbor Laboratory Press.
Figures


References
-
- Ardiel EL, Rankin CH. 2010. An elegant mind: learning and memory in Caenorhabditis elegans. Learn Mem 17: 191–201. - PubMed
-
- Bargmann CI, Thomas JH, Horvitz HR. 1990. Chemosensory cell function in the behavior and development of Caenorhabditis elegans. Cold Spring Harb Symp Quant Biol 55: 529–538. - PubMed
-
- Bettinger JC, McIntire SL. 2004. State-dependency in C. elegans. Genes Brain Behav 3: 266–272. - PubMed
-
- Braff DL, Grillon C, Geyer MA. 1992. Gating and habituation of the startle reflex in schizophrenic patients. Arch Gen Psychiatry 49: 206–215. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous