

Chromatin determinants of the inner-centromere rely on replication factors with functions that impart cohesion
- PMID: 27636994
- PMCID: PMC5356530
- DOI: 10.18632/oncotarget.11982
Chromatin determinants of the inner-centromere rely on replication factors with functions that impart cohesion
Abstract
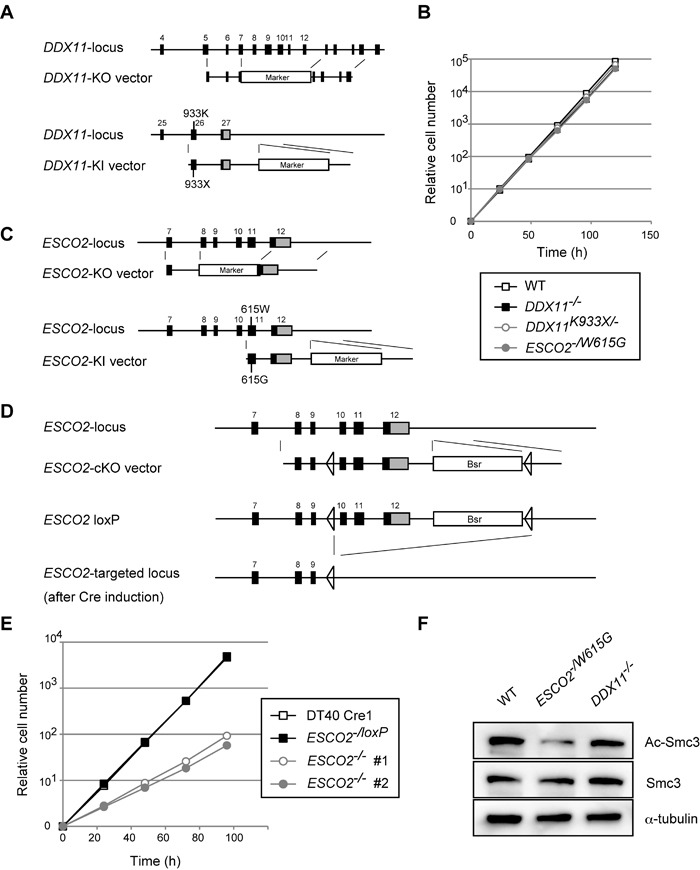
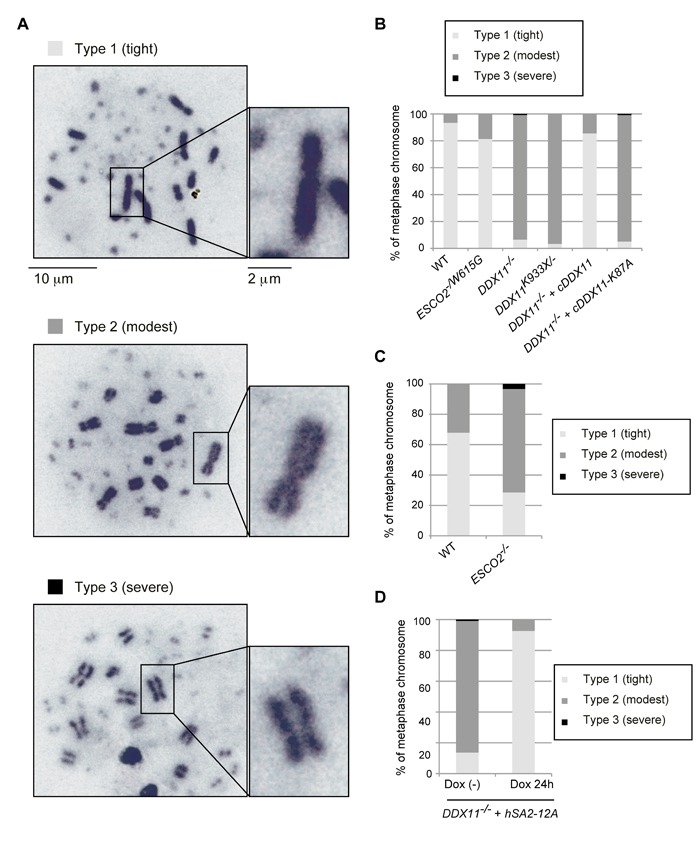
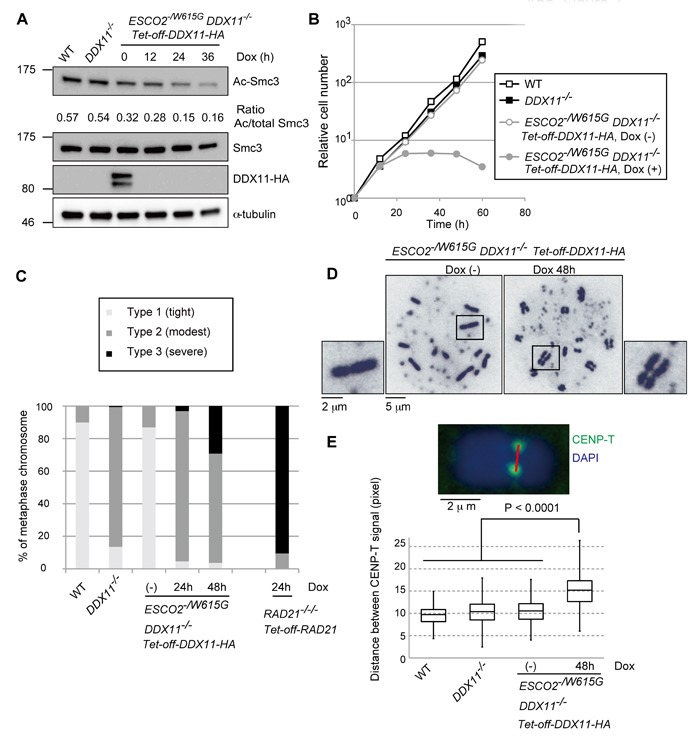
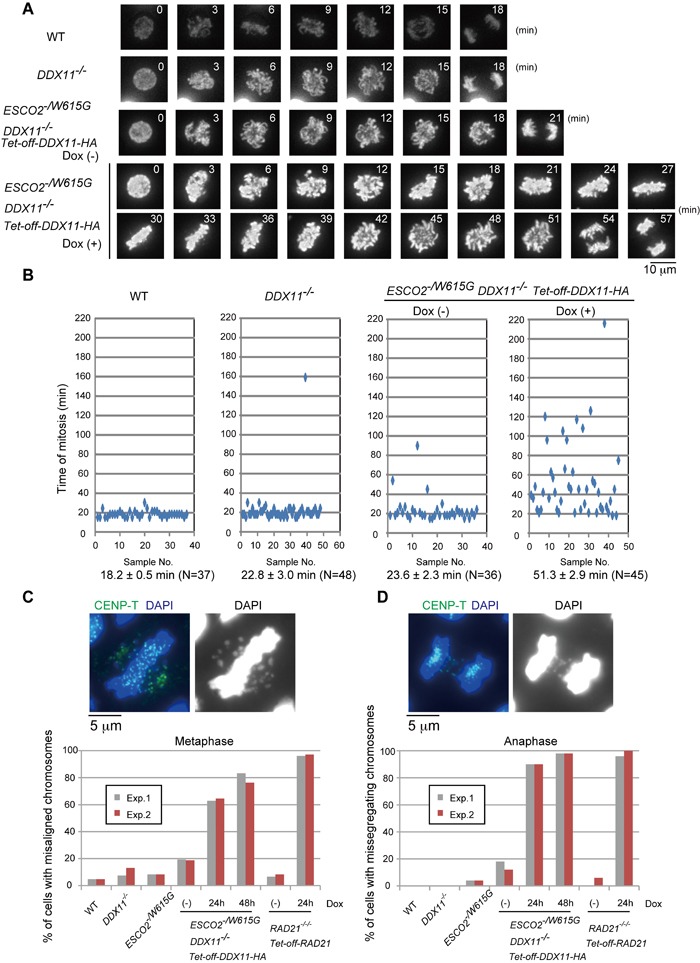
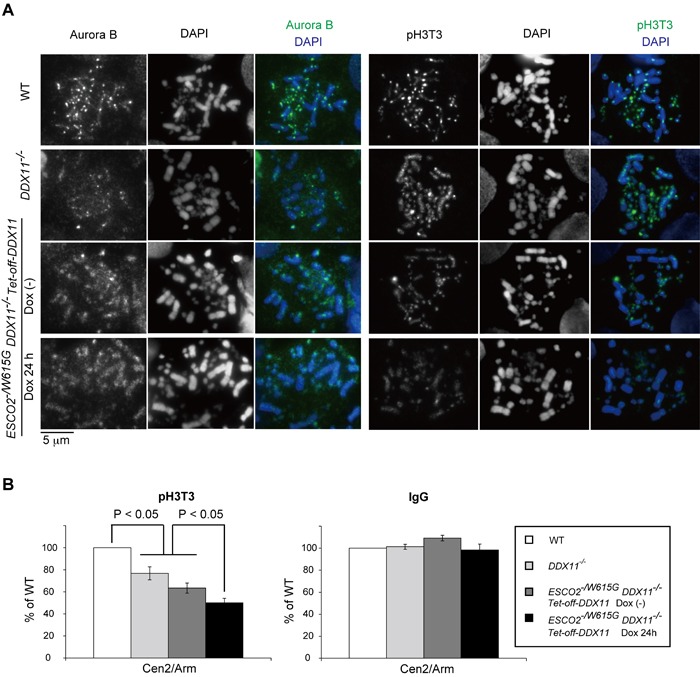
Replication fork-associated factors promote genome integrity and protect against cancer. Mutations in the DDX11 helicase and the ESCO2 acetyltransferase also cause related developmental disorders classified as cohesinopathies. Here we generated vertebrate model cell lines of these disorders and cohesinopathies-related genes. We found that vertebrate DDX11 and Tim-Tipin are individually needed to compensate for ESCO2 loss in chromosome segregation, with DDX11 also playing complementary roles with ESCO2 in centromeric cohesion. Our study reveals that overt centromeric cohesion loss does not necessarily precede chromosome missegregation, while both these problems correlate with, and possibly originate from, inner-centromere defects involving reduced phosphorylation of histone H3T3 (pH3T3) in the region. Interestingly, the mitotic pH3T3 mark was defective in all analyzed replication-related mutants with functions in cohesion. The results pinpoint mitotic pH3T3 as a postreplicative chromatin mark that is sensitive to replication stress and conducts with different kinetics to robust centromeric cohesion and correct chromosome segregation.
Keywords: DDX11; Tim-Tipin; inner-centromere; replication stress; sister chromatid cohesion; Chromosome Section.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures

References
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
