

Multi-tiered Reorganization of the Genome during B Cell Affinity Maturation Anchored by a Germinal Center-Specific Locus Control Region
- PMID: 27637145
- PMCID: PMC5033726
- DOI: 10.1016/j.immuni.2016.08.012
Multi-tiered Reorganization of the Genome during B Cell Affinity Maturation Anchored by a Germinal Center-Specific Locus Control Region
Abstract
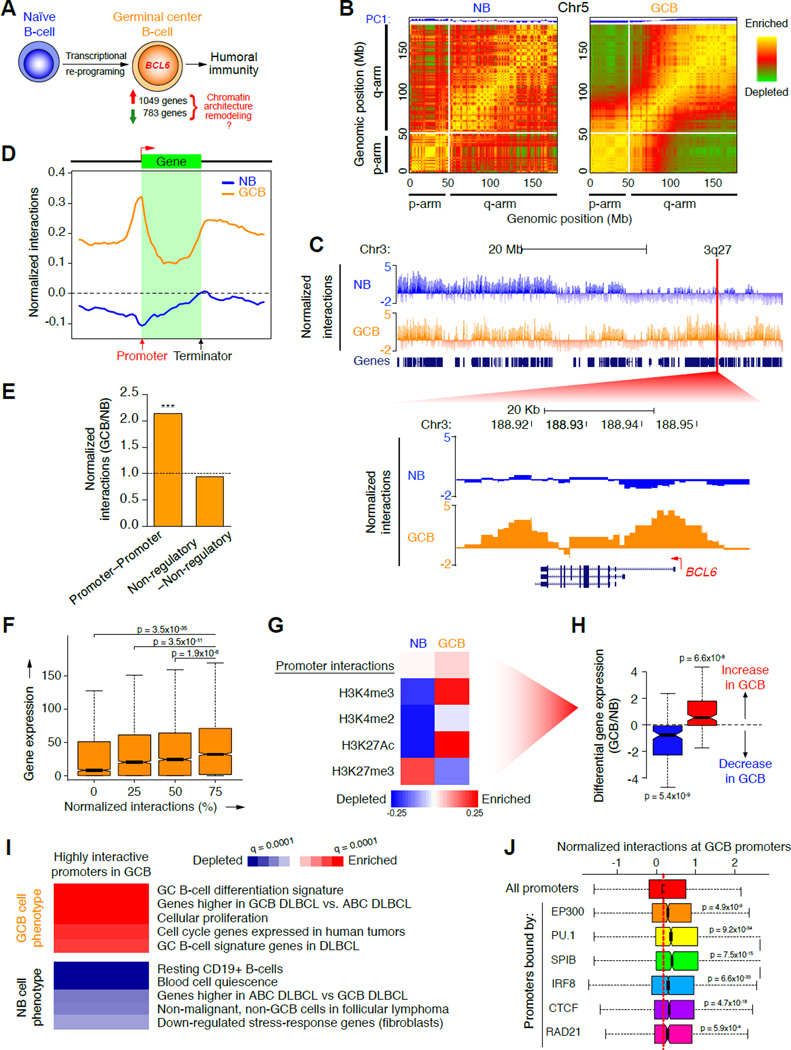
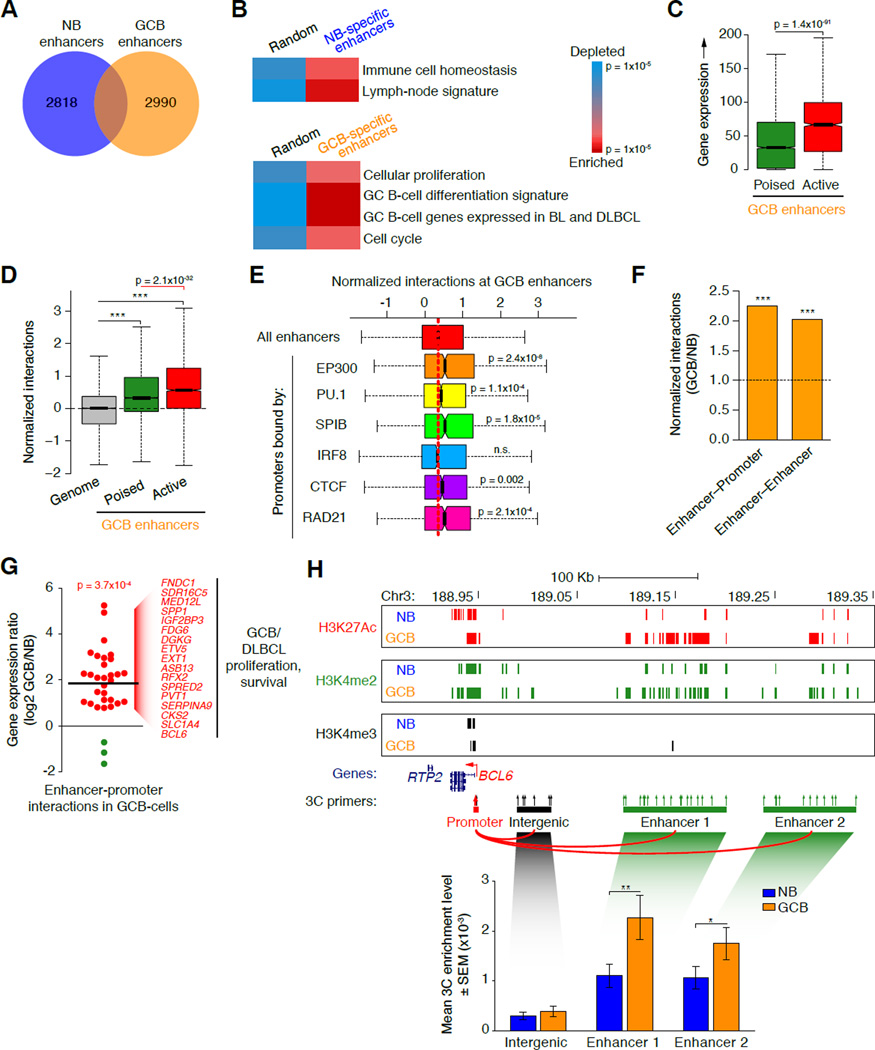
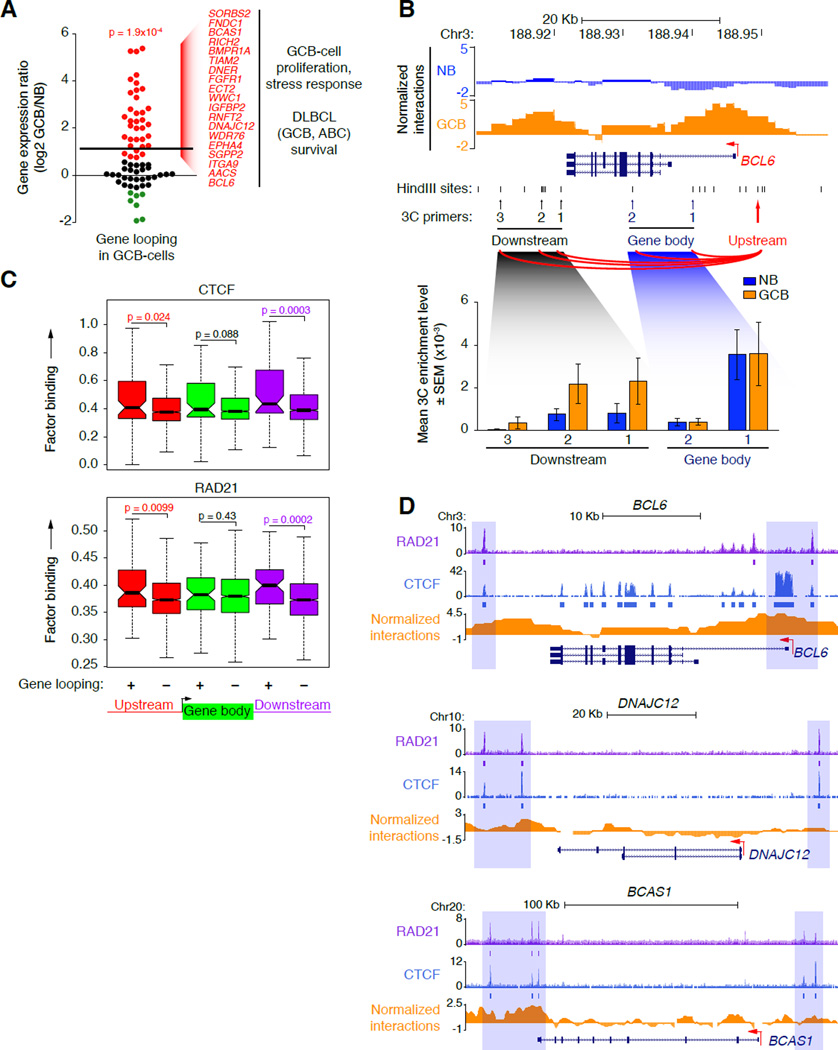
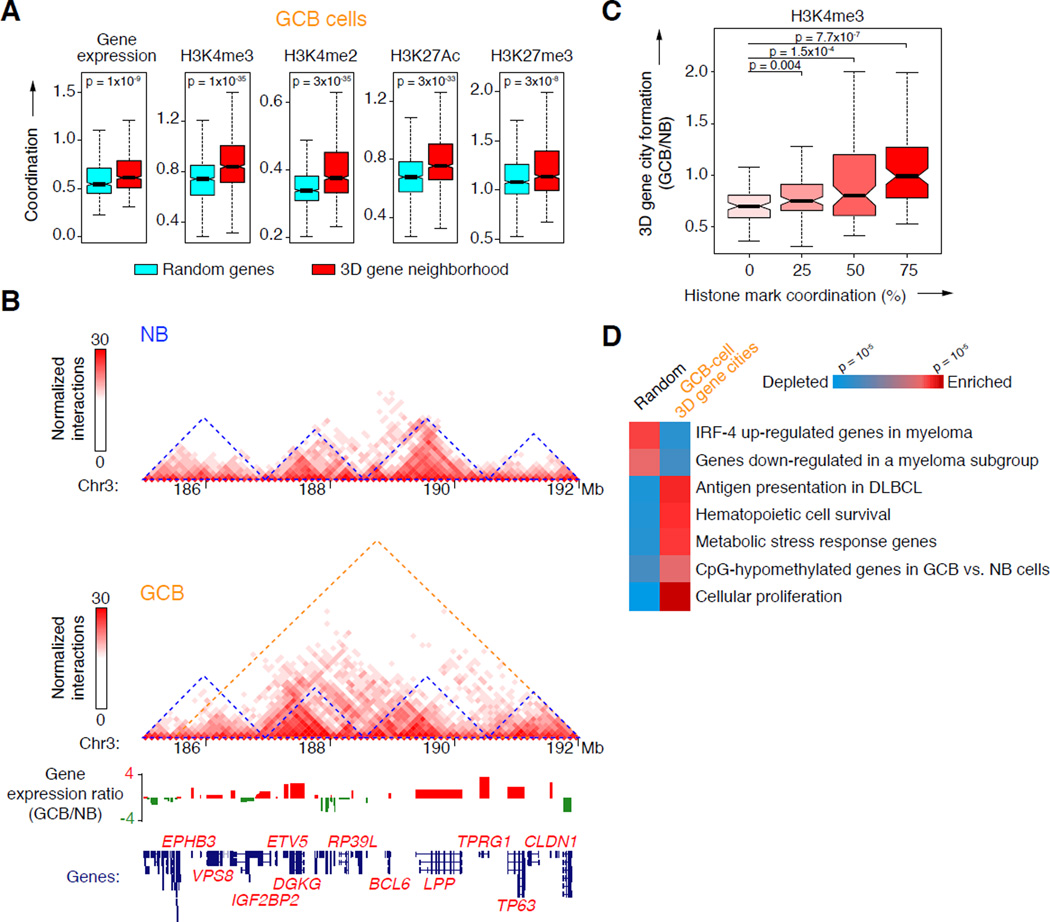
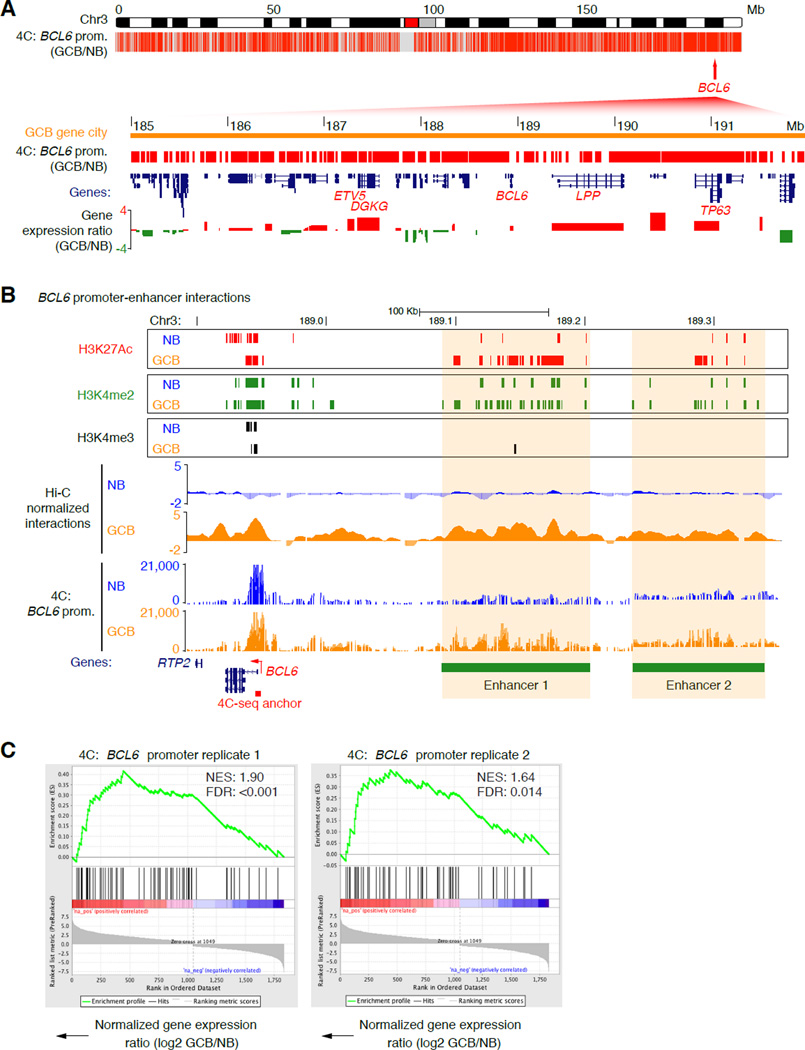
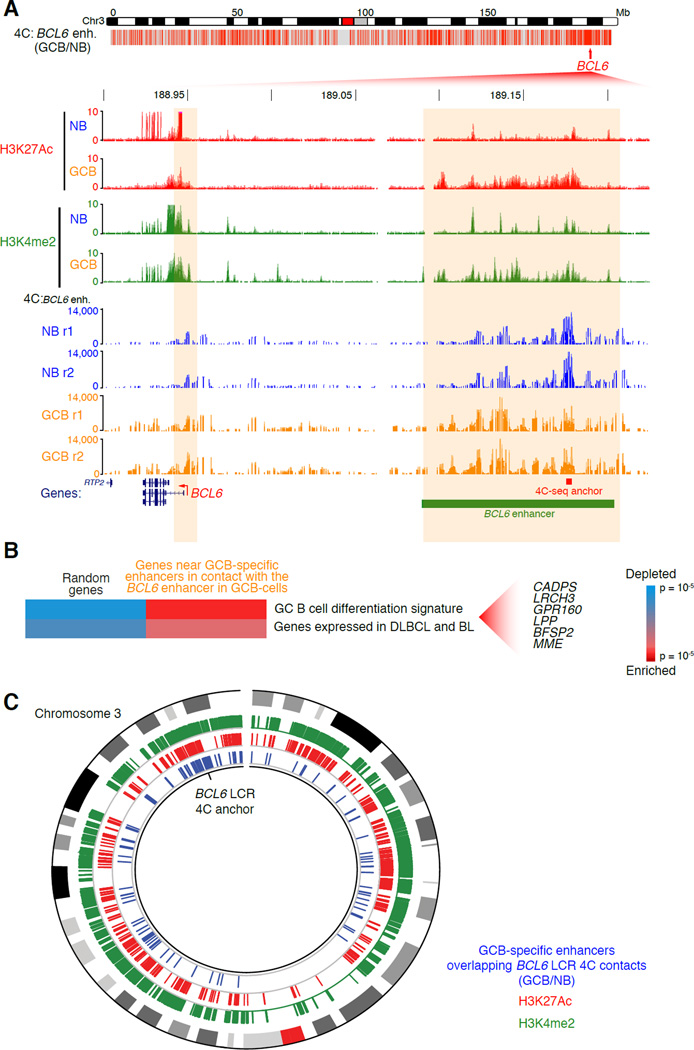
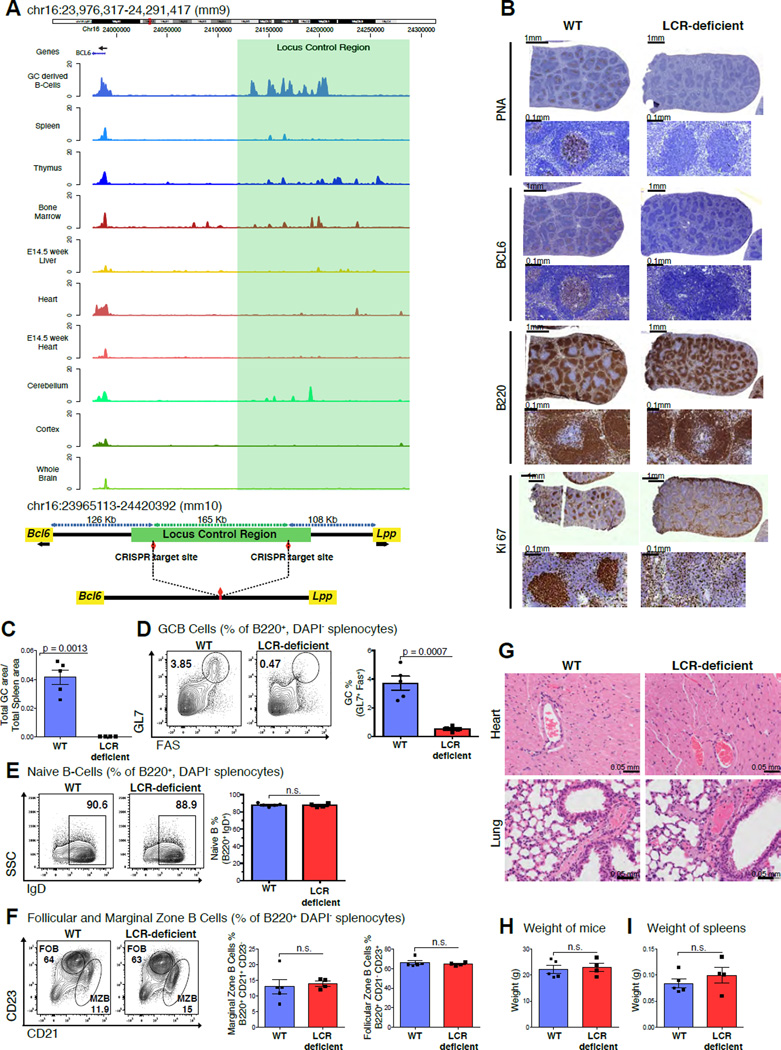
During the humoral immune response, B cells undergo a dramatic change in phenotype to enable antibody affinity maturation in germinal centers (GCs). Using genome-wide chromosomal conformation capture (Hi-C), we found that GC B cells undergo massive reorganization of the genomic architecture that encodes the GC B cell transcriptome. Coordinate expression of genes that specify the GC B cell phenotype-most prominently BCL6-was achieved through a multilayered chromatin reorganization process involving (1) increased promoter connectivity, (2) formation of enhancer networks, (3) 5' to 3' gene looping, and (4) merging of gene neighborhoods that share active epigenetic marks. BCL6 was an anchor point for the formation of GC-specific gene and enhancer loops on chromosome 3. Deletion of a GC-specific, highly interactive locus control region upstream of Bcl6 abrogated GC formation in mice. Thus, large-scale and multi-tiered genomic three-dimensional reorganization is required for coordinate expression of phenotype-driving gene sets that determine the unique characteristics of GC B cells.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Looping around Bcl6 in Germinal Center to Sharpen B Cell Immunity.Immunity. 2016 Sep 20;45(3):459-461. doi: 10.1016/j.immuni.2016.08.020. Immunity. 2016. PMID: 27653595 Free PMC article.
-
B cells: Genomic reorganization in germinal centres.Nat Rev Immunol. 2016 Sep 27;16(10):594-5. doi: 10.1038/nri.2016.108. Nat Rev Immunol. 2016. PMID: 27670367 No abstract available.
References
-
- Bai Y, Srinivasan L, Perkins L, Atchison ML. Protein acetylation regulates both PU.1 transactivation and Ig kappa 3' enhancer activity. J Immunol. 2005;175:5160–5169. - PubMed
-
- Bickmore WA, van Steensel B. Genome architecture: domain organization of interphase chromosomes. Cell. 2013;152:1270–1284. - PubMed
-
- Cantone I, Fisher AG. Epigenetic programming and reprogramming during development. Nat Struct Mol Biol. 2013;20:282–289. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
