

Unsupervised High-Dimensional Analysis Aligns Dendritic Cells across Tissues and Species
- PMID: 27637149
- PMCID: PMC5040826
- DOI: 10.1016/j.immuni.2016.08.015
Unsupervised High-Dimensional Analysis Aligns Dendritic Cells across Tissues and Species
Abstract
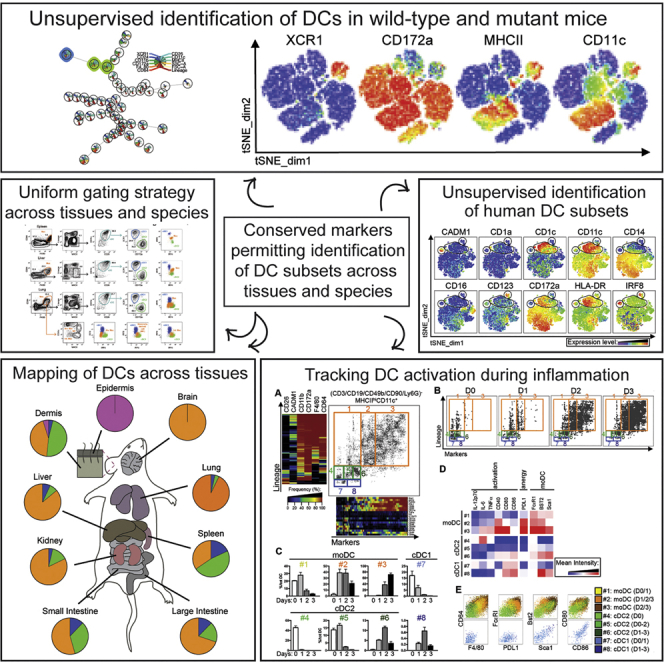
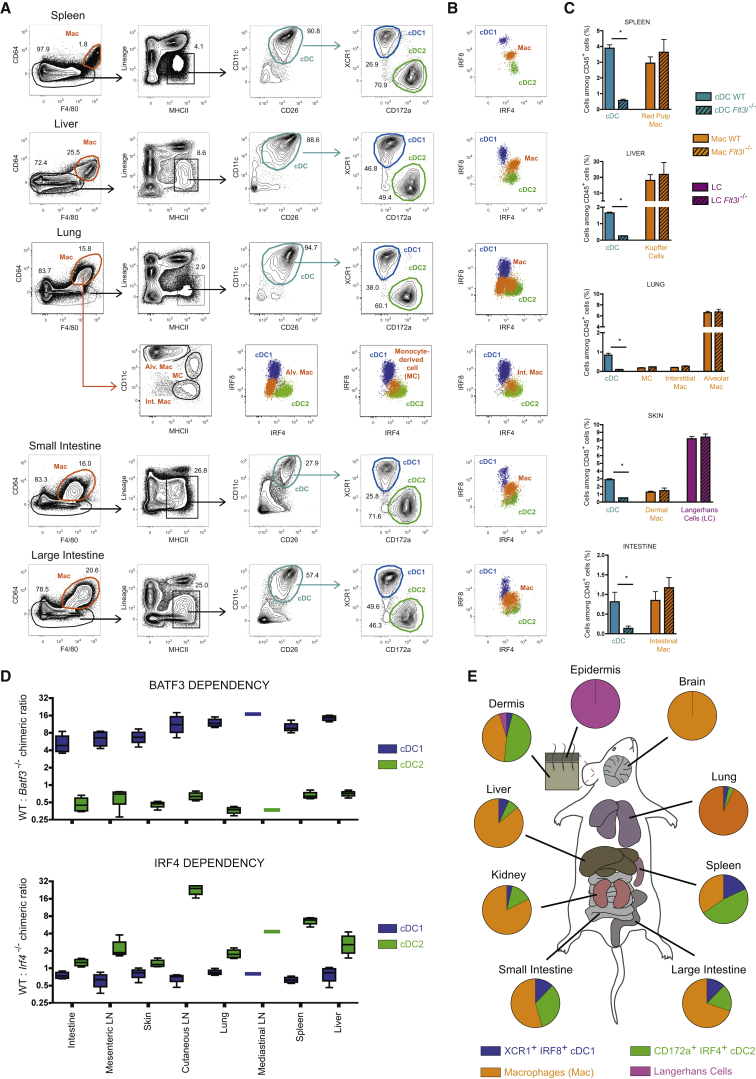
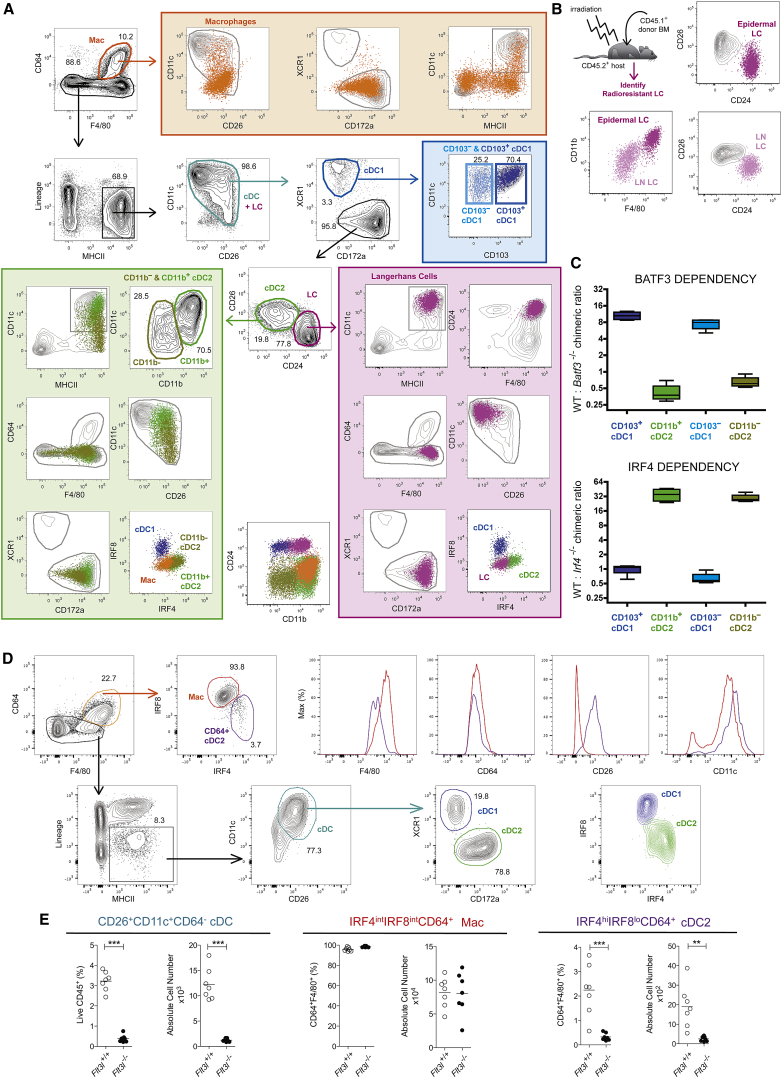
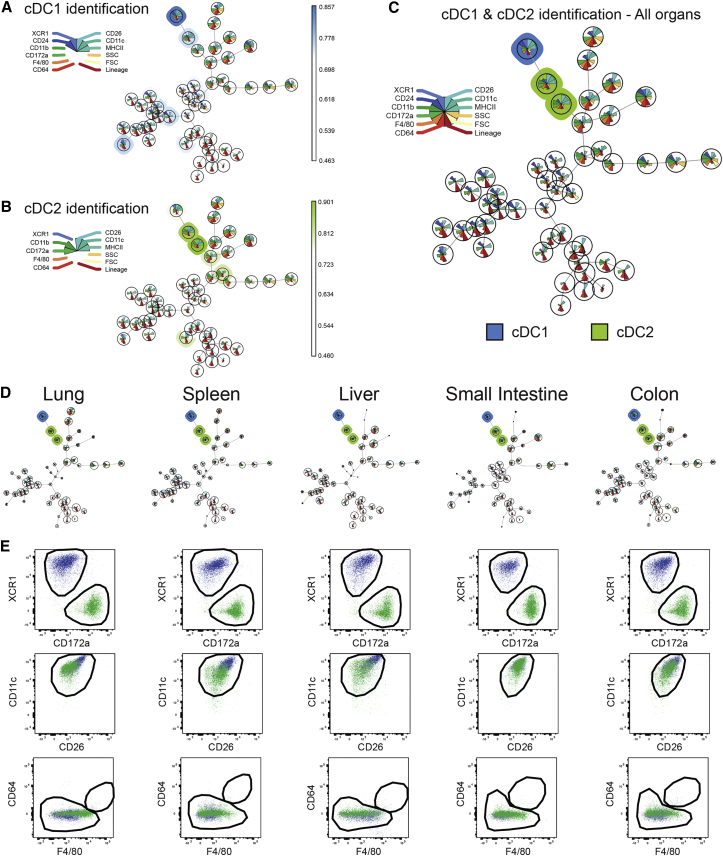
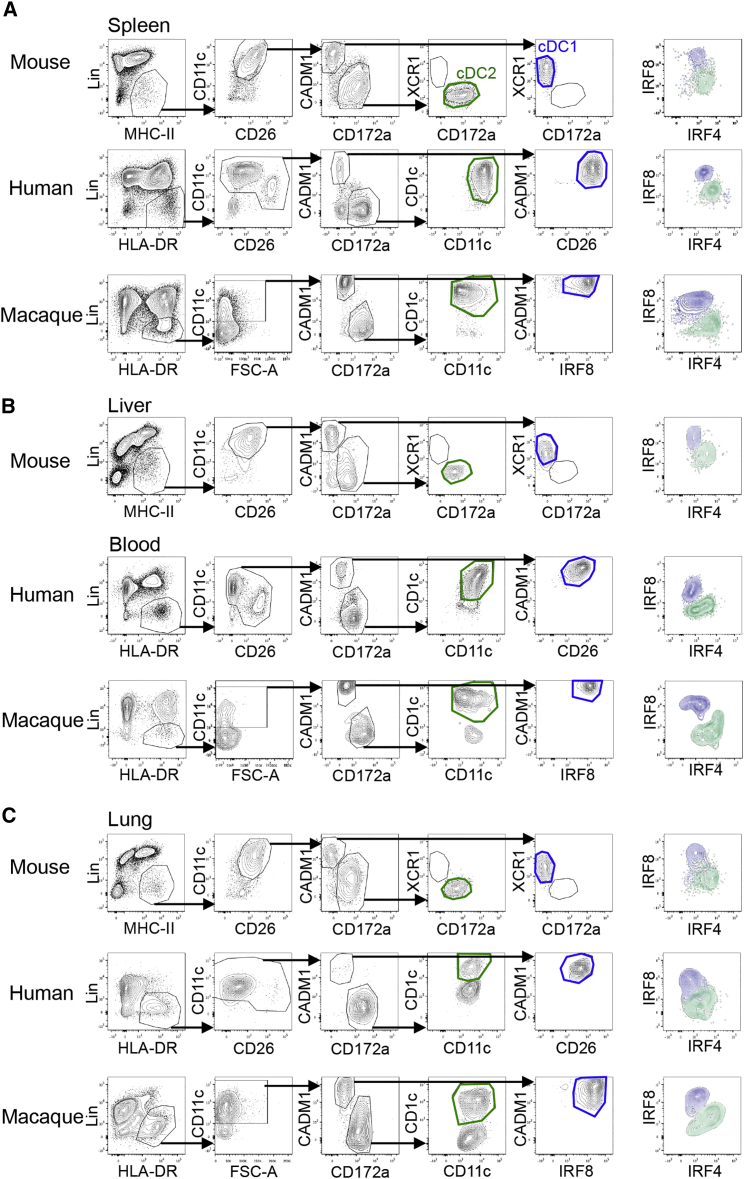
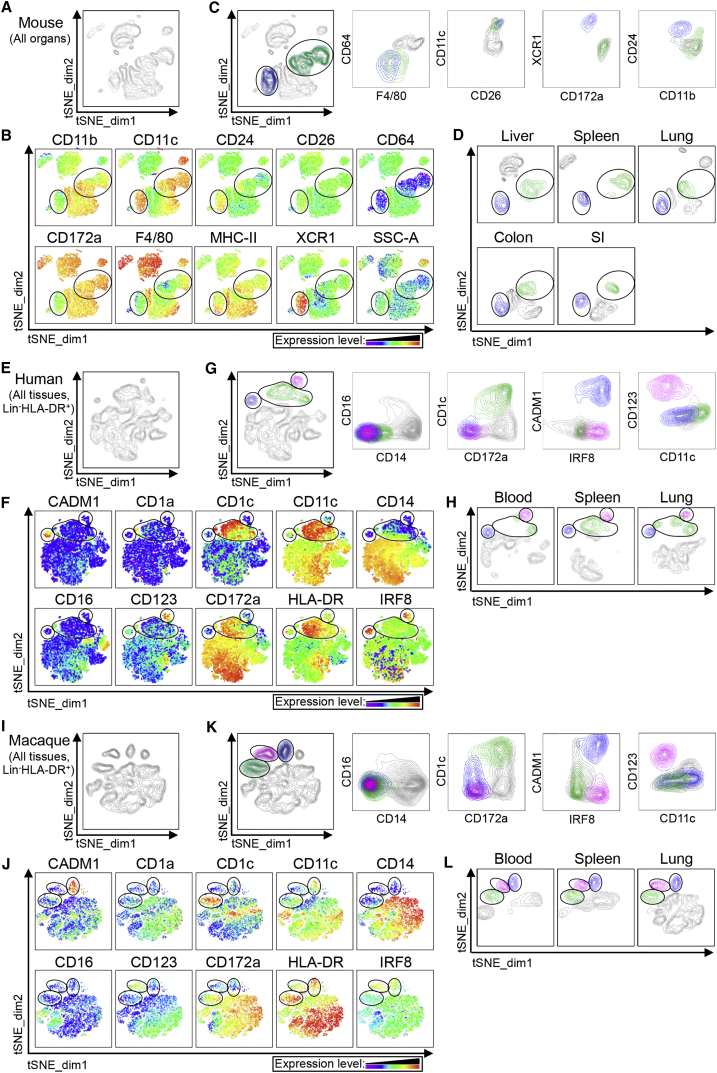
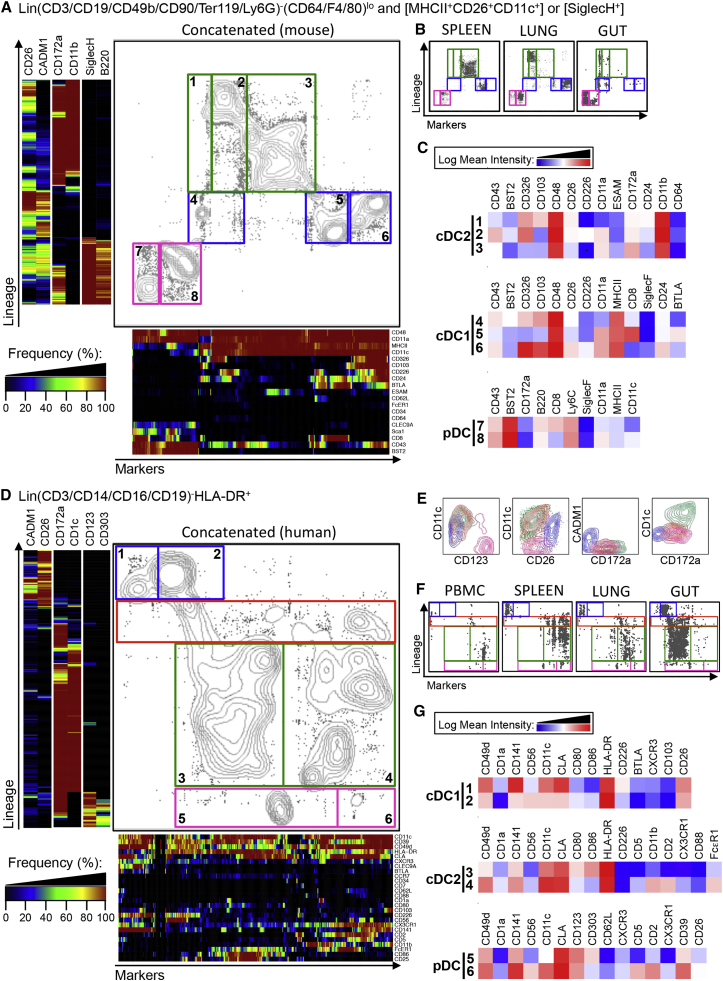
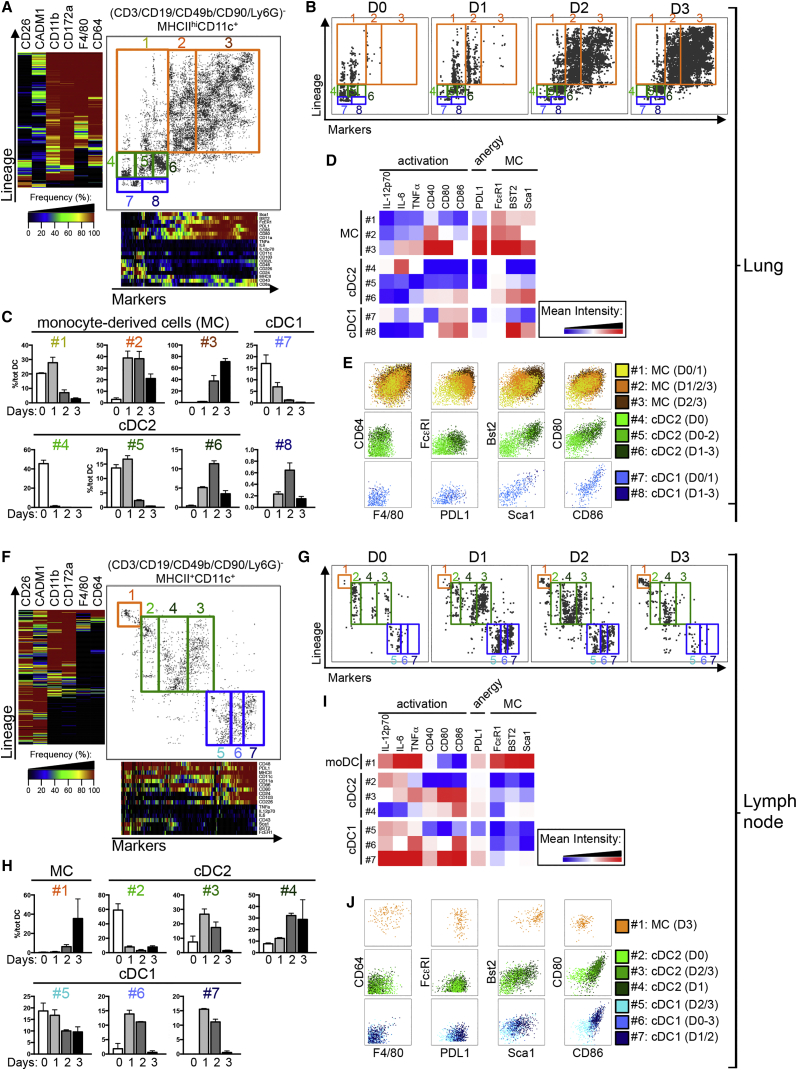
Dendritic cells (DCs) are professional antigen-presenting cells that hold great therapeutic potential. Multiple DC subsets have been described, and it remains challenging to align them across tissues and species to analyze their function in the absence of macrophage contamination. Here, we provide and validate a universal toolbox for the automated identification of DCs through unsupervised analysis of conventional flow cytometry and mass cytometry data obtained from multiple mouse, macaque, and human tissues. The use of a minimal set of lineage-imprinted markers was sufficient to subdivide DCs into conventional type 1 (cDC1s), conventional type 2 (cDC2s), and plasmacytoid DCs (pDCs) across tissues and species. This way, a large number of additional markers can still be used to further characterize the heterogeneity of DCs across tissues and during inflammation. This framework represents the way forward to a universal, high-throughput, and standardized analysis of DC populations from mutant mice and human patients.
Copyright © 2016 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
Comment in
-
Dendritic cells: Sorting, sorted!Nat Rev Immunol. 2016 Nov;16(11):657. doi: 10.1038/nri.2016.115. Epub 2016 Oct 17. Nat Rev Immunol. 2016. PMID: 27748396 No abstract available.
References
-
- Bain C.C., Scott C.L., Uronen-Hansson H., Gudjonsson S., Jansson O., Grip O., Guilliams M., Malissen B., Agace W.W., Mowat A.M. Resident and pro-inflammatory macrophages in the colon represent alternative context-dependent fates of the same Ly6C(hi) monocyte precursors. Mucosal Immunol. 2012;6:498–510. - PMC - PubMed
-
- Becher B., Schlitzer A., Chen J., Mair F., Sumatoh H.R., Teng K.W., Low D., Ruedl C., Riccardi-Castagnoli P., Poidinger M. High-dimensional analysis of the murine myeloid cell system. Nat. Immunol. 2014;15:1181–1189. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
