

Cortical dynamics during cell motility are regulated by CRL3(KLHL21) E3 ubiquitin ligase
- PMID: 27641145
- PMCID: PMC5031805
- DOI: 10.1038/ncomms12810
Cortical dynamics during cell motility are regulated by CRL3(KLHL21) E3 ubiquitin ligase
Erratum in
-
Corrigendum: Cortical dynamics during cell motility are regulated by CRL3KLHL21 E3 ubiquitin ligase.Nat Commun. 2016 Oct 31;7:13487. doi: 10.1038/ncomms13487. Nat Commun. 2016. PMID: 27796354 Free PMC article. No abstract available.
Abstract
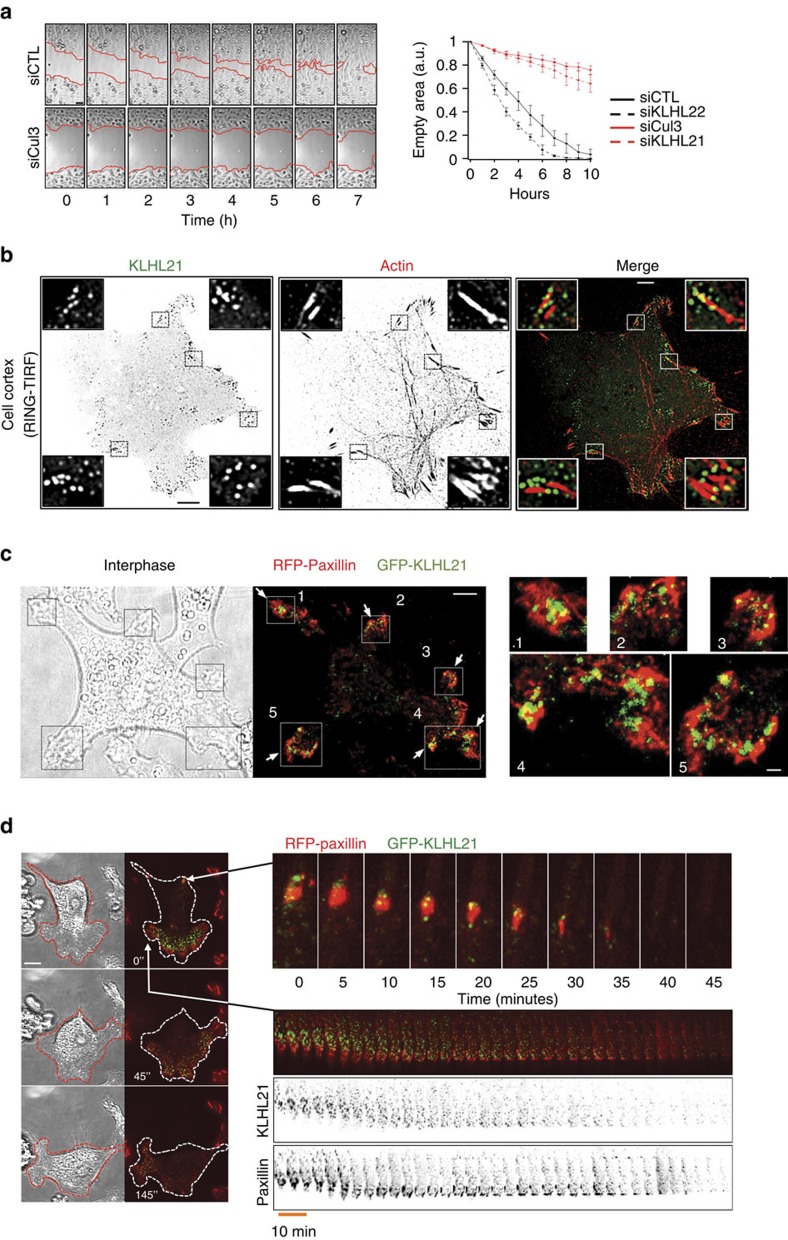
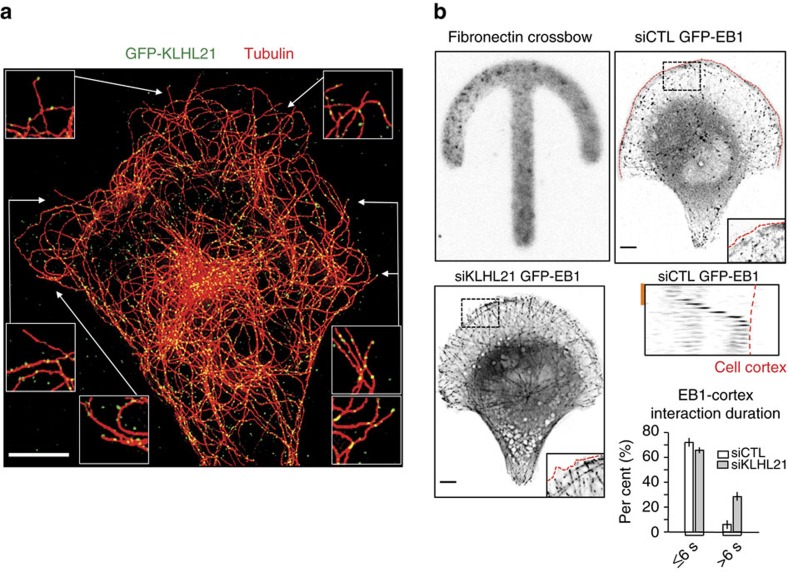
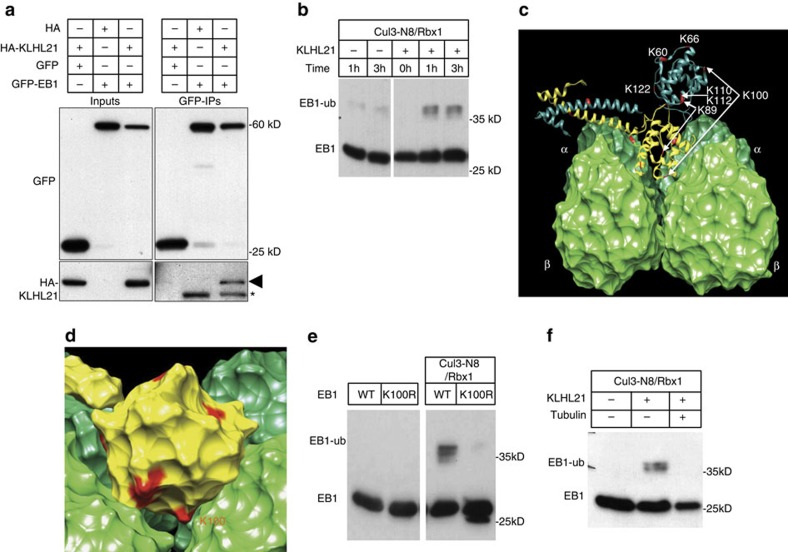
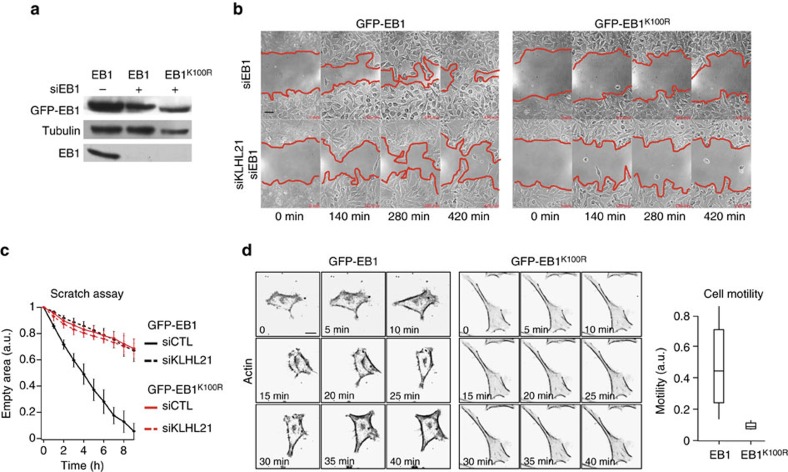
Directed cell movement involves spatial and temporal regulation of the cortical microtubule (Mt) and actin networks to allow focal adhesions (FAs) to assemble at the cell front and disassemble at the rear. Mts are known to associate with FAs, but the mechanisms coordinating their dynamic interactions remain unknown. Here we show that the CRL3(KLHL21) E3 ubiquitin ligase promotes cell migration by controlling Mt and FA dynamics at the cell cortex. Indeed, KLHL21 localizes to FA structures preferentially at the leading edge, and in complex with Cul3, ubiquitylates EB1 within its microtubule-interacting CH-domain. Cells lacking CRL3(KLHL21) activity or expressing a non-ubiquitylatable EB1 mutant protein are unable to migrate and exhibit strong defects in FA dynamics, lamellipodia formation and cortical plasticity. Our study thus reveals an important mechanism to regulate cortical dynamics during cell migration that involves ubiquitylation of EB1 at focal adhesions.
Figures
Similar articles
-
HECTD1 controls the protein level of IQGAP1 to regulate the dynamics of adhesive structures.Cell Commun Signal. 2017 Jan 5;15(1):2. doi: 10.1186/s12964-016-0156-8. Cell Commun Signal. 2017. PMID: 28073378 Free PMC article.
-
Proto-Oncogenic Src Phosphorylates EB1 to Regulate the Microtubule-Focal Adhesion Crosstalk and Stimulate Cell Migration.Theranostics. 2016 Sep 12;6(12):2129-2140. doi: 10.7150/thno.16356. eCollection 2016. Theranostics. 2016. PMID: 27698945 Free PMC article.
-
Ubiquitylation of phosphatidylinositol 4-phosphate 5-kinase type I γ by HECTD1 regulates focal adhesion dynamics and cell migration.J Cell Sci. 2013 Jun 15;126(Pt 12):2617-28. doi: 10.1242/jcs.117044. Epub 2013 Apr 9. J Cell Sci. 2013. PMID: 23572508 Free PMC article.
-
Asymmetric focal adhesion disassembly in motile cells.Curr Opin Cell Biol. 2008 Feb;20(1):85-90. doi: 10.1016/j.ceb.2007.10.009. Curr Opin Cell Biol. 2008. PMID: 18083360 Review.
-
Assembly and mechanosensory function of focal adhesions: experiments and models.Eur J Cell Biol. 2006 Apr;85(3-4):165-73. doi: 10.1016/j.ejcb.2005.11.001. Epub 2005 Dec 19. Eur J Cell Biol. 2006. PMID: 16360240 Review.
Cited by
-
Modulation of Abiotic Stress Responses in Rice by E3-Ubiquitin Ligases: A Promising Way to Develop Stress-Tolerant Crops.Front Plant Sci. 2021 Mar 23;12:640193. doi: 10.3389/fpls.2021.640193. eCollection 2021. Front Plant Sci. 2021. PMID: 33833769 Free PMC article. Review.
-
The roles of KLHL family members in human cancers.Am J Cancer Res. 2022 Nov 15;12(11):5105-5139. eCollection 2022. Am J Cancer Res. 2022. PMID: 36504893 Free PMC article. Review.
-
Functional analysis of Cullin 3 E3 ligases in tumorigenesis.Biochim Biophys Acta Rev Cancer. 2018 Jan;1869(1):11-28. doi: 10.1016/j.bbcan.2017.11.001. Epub 2017 Nov 8. Biochim Biophys Acta Rev Cancer. 2018. PMID: 29128526 Free PMC article. Review.
-
KLHL5 knockdown increases cellular sensitivity to anticancer drugs.Oncotarget. 2018 Dec 21;9(100):37429-37438. doi: 10.18632/oncotarget.26462. eCollection 2018 Dec 21. Oncotarget. 2018. PMID: 30647843 Free PMC article.
-
Cullin 3-Based Ubiquitin Ligases as Master Regulators of Mammalian Cell Differentiation.Trends Biochem Sci. 2018 Feb;43(2):95-107. doi: 10.1016/j.tibs.2017.11.010. Epub 2017 Dec 14. Trends Biochem Sci. 2018. PMID: 29249570 Free PMC article. Review.
References
-
- Akhshi T. K., Wernike D. & Piekny A. Microtubules and actin crosstalk in cell migration and division. Cytoskeleton (Hoboken) 71, 1–23 (2014). - PubMed
-
- Vasiliev J. M. et al.. Effect of colcemid on the locomotory behaviour of fibroblasts. J. Embryol. Exp. Morphol. 24, 625–640 (1970). - PubMed
-
- Mitchison T. & Kirschner M. Dynamic instability of microtubule growth. Nature 312, 237–242 (1984). - PubMed
-
- Jiang K. & Akhmanova A. S. Microtubule tip-interacting proteins: a view from both ends. Curr. Opin. Cell Biol. 23, 94–101 (2011). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
