

Cellular adaptation to biomechanical stress across length scales in tissue homeostasis and disease
- PMID: 27641825
- PMCID: PMC5352546
- DOI: 10.1016/j.semcdb.2016.09.004
Cellular adaptation to biomechanical stress across length scales in tissue homeostasis and disease
Abstract
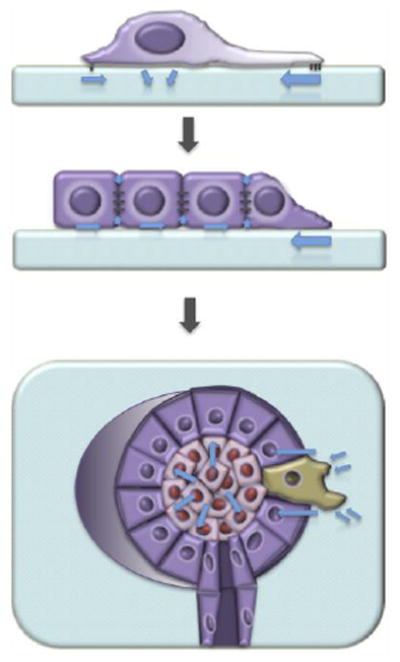
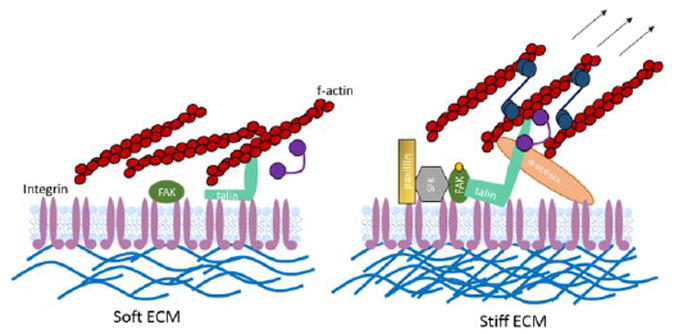
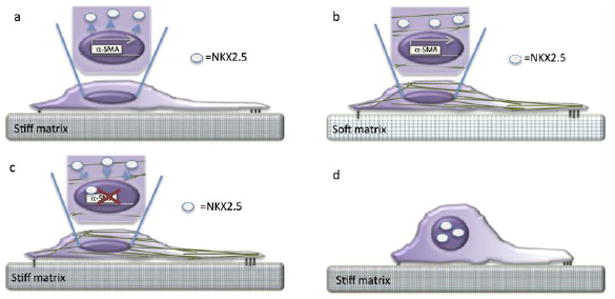
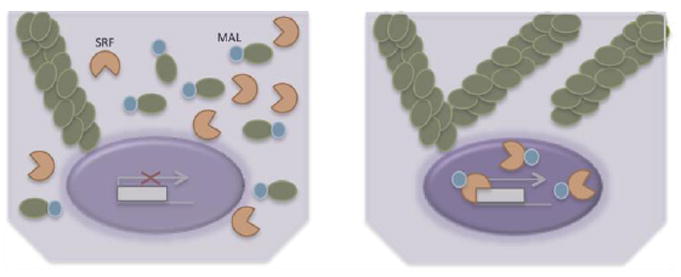
Human tissues are remarkably adaptable and robust, harboring the collective ability to detect and respond to external stresses while maintaining tissue integrity. Following injury, many tissues have the capacity to repair the damage - and restore form and function - by deploying cellular and molecular mechanisms reminiscent of developmental programs. Indeed, it is increasingly clear that cancer and chronic conditions that develop with age arise as a result of cells and tissues re-implementing and deregulating a selection of developmental programs. Therefore, understanding the fundamental molecular mechanisms that drive cell and tissue responses is a necessity when designing therapies to treat human conditions. Extracellular matrix stiffness synergizes with chemical cues to drive single cell and collective cell behavior in culture and acts to establish and maintain tissue homeostasis in the body. This review will highlight recent advances that elucidate the impact of matrix mechanics on cell behavior and fate across these length scales during times of homeostasis and in disease states.
Keywords: Actin-myosin contractility; Biomechanics; Cancer; Cell contractility; Development; EMT; Intracellular tension; Matrix stiffness; Mechanical force; Mechanical memory; Mechanosensing; Mechanotransduction; Tissue homeostasis; Tissue tension.
Copyright © 2016 Elsevier Ltd. All rights reserved.
Figures
References
-
- HUXLEY AF, NIEDERGERKE R. Structural changes in muscle during contraction; interference microscopy of living muscle fibres. [November 18, 2015];Nature. 1954 173:971–3. http://www.ncbi.nlm.nih.gov/pubmed/13165697. - PubMed
-
- HUXLEY H, HANSON J. Changes in the cross-striations of muscle during contraction and stretch and their structural interpretation. [October 16, 2015];Nature. 1954 173:973–6. http://www.ncbi.nlm.nih.gov/pubmed/13165698. - PubMed
-
- Abercrombie M, Heaysman JE, Pegrum SM. The locomotion of fibroblasts in culture. IV. Electron microscopy of the leading lamella. [February 19, 2016];Exp Cell Res. 1971 67:359–67. http://www.ncbi.nlm.nih.gov/pubmed/5097522. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
