

Dysferlin-mediated phosphatidylserine sorting engages macrophages in sarcolemma repair
- PMID: 27641898
- PMCID: PMC5031802
- DOI: 10.1038/ncomms12875
Dysferlin-mediated phosphatidylserine sorting engages macrophages in sarcolemma repair
Abstract
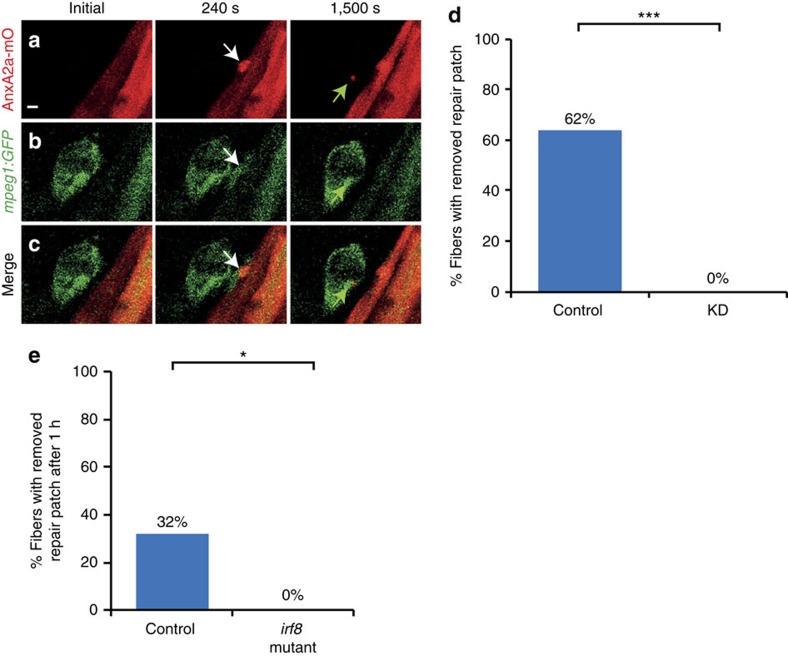
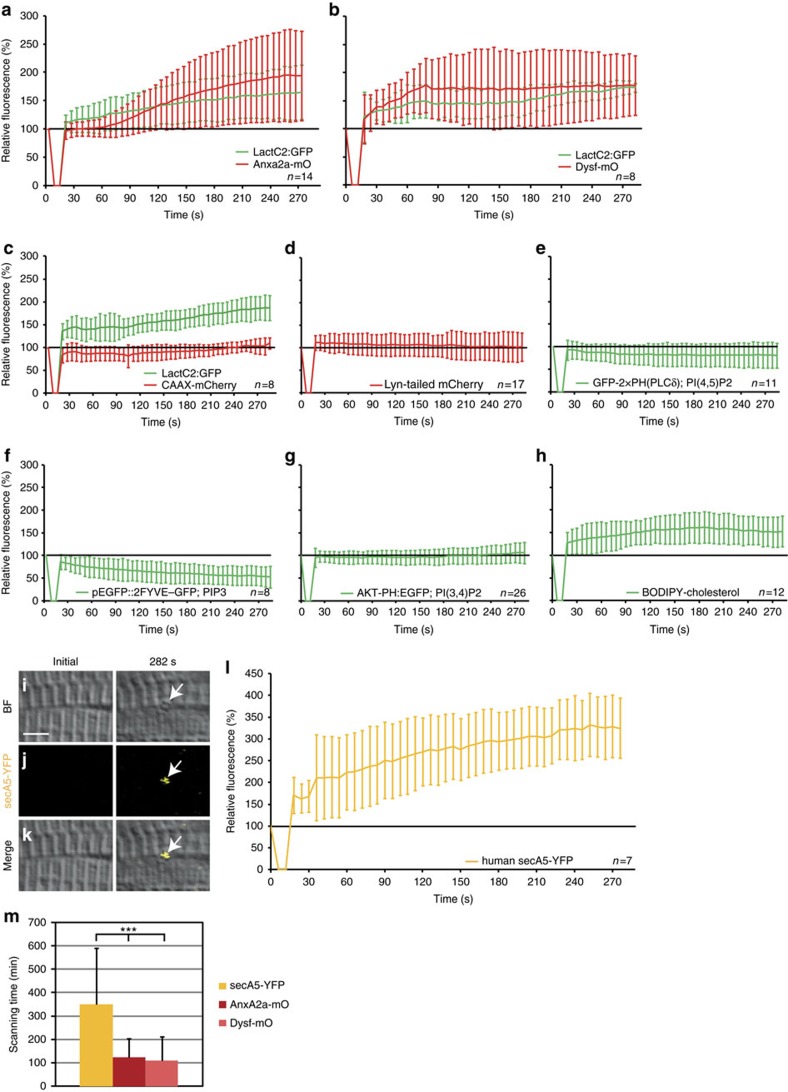
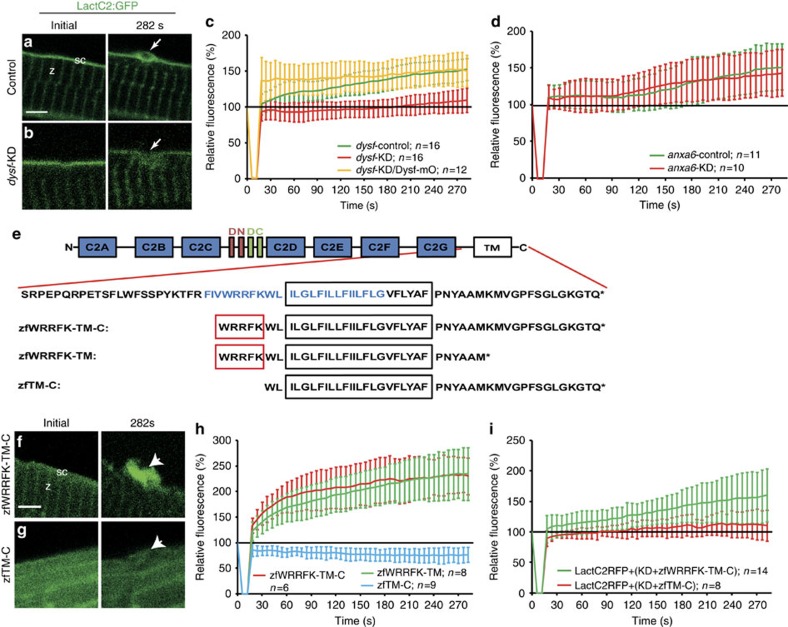
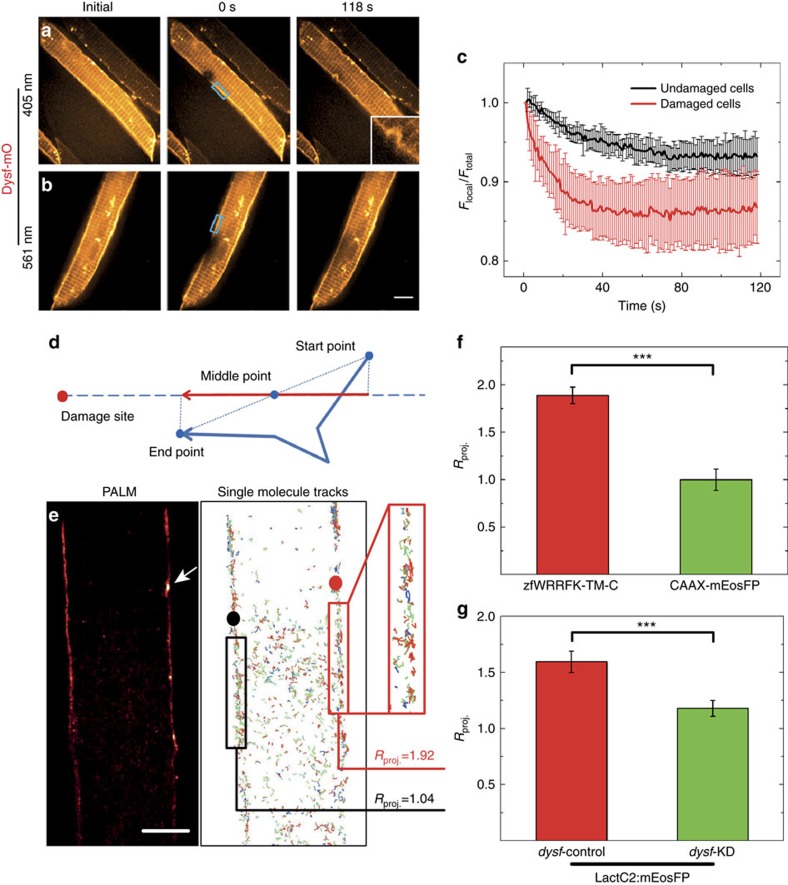
Failure to repair the sarcolemma leads to muscle cell death, depletion of stem cells and myopathy. Hence, membrane lesions are instantly sealed by a repair patch consisting of lipids and proteins. It has remained elusive how this patch is removed to restore cell membrane integrity. Here we examine sarcolemmal repair in live zebrafish embryos by real-time imaging. Macrophages remove the patch. Phosphatidylserine (PS), an 'eat-me' signal for macrophages, is rapidly sorted from adjacent sarcolemma to the repair patch in a Dysferlin (Dysf) dependent process in zebrafish and human cells. A previously unrecognized arginine-rich motif in Dysf is crucial for PS accumulation. It carries mutations in patients presenting with limb-girdle muscular dystrophy 2B. This underscores the relevance of this sequence and uncovers a novel pathophysiological mechanism underlying this class of myopathies. Our data show that membrane repair is a multi-tiered process involving immediate, cell-intrinsic mechanisms as well as myofiber/macrophage interactions.
Figures
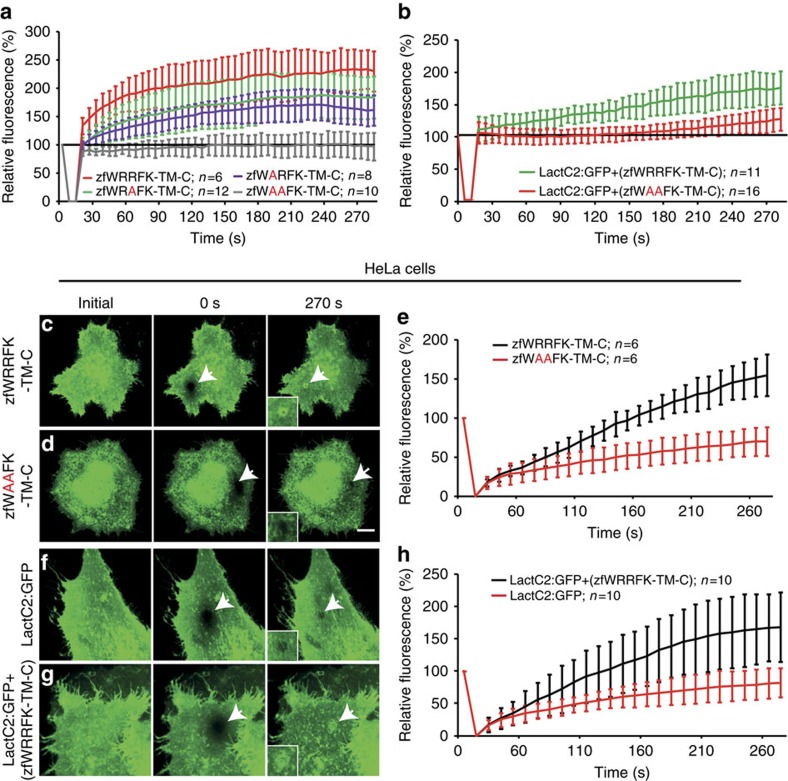
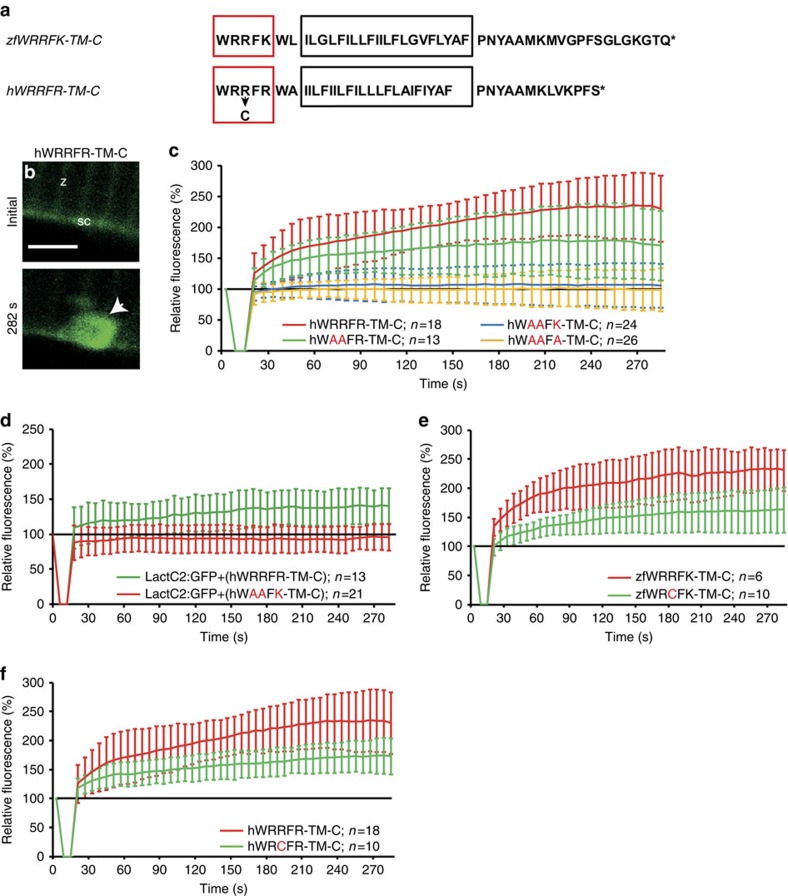
References
-
- Bansal D. et al.. Defective membrane repair in dysferlin-deficient muscular dystrophy. Nature 423, 168–172 (2003). - PubMed
-
- Bashir R. et al.. A gene related to Caenorhabditis elegans spermatogenesis factor fer-1 is mutated in limb-girdle muscular dystrophy type 2B. Nat. Genet. 20, 37–42 (1998). - PubMed
-
- Liu J. et al.. Dysferlin, a novel skeletal muscle gene, is mutated in Miyoshi myopathy and limb girdle muscular dystrophy. Nat. Genet. 20, 31–36 (1998). - PubMed
-
- Therrien C., Di Fulvio S., Pickles S. & Sinnreich M. Characterization of lipid binding specificities of dysferlin C2 domains reveals novel interactions with phosphoinositides. Biochemistry 48, 2377–2384 (2009). - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
