

Coordinated nuclease activities counteract Ku at single-ended DNA double-strand breaks
- PMID: 27641979
- PMCID: PMC5031800
- DOI: 10.1038/ncomms12889
Coordinated nuclease activities counteract Ku at single-ended DNA double-strand breaks
Erratum in
-
Corrigendum: Coordinated nuclease activities counteract Ku at single-ended DNA double-strand breaks.Nat Commun. 2017 Jun 13;8:15917. doi: 10.1038/ncomms15917. Nat Commun. 2017. PMID: 28607502 Free PMC article.
Abstract
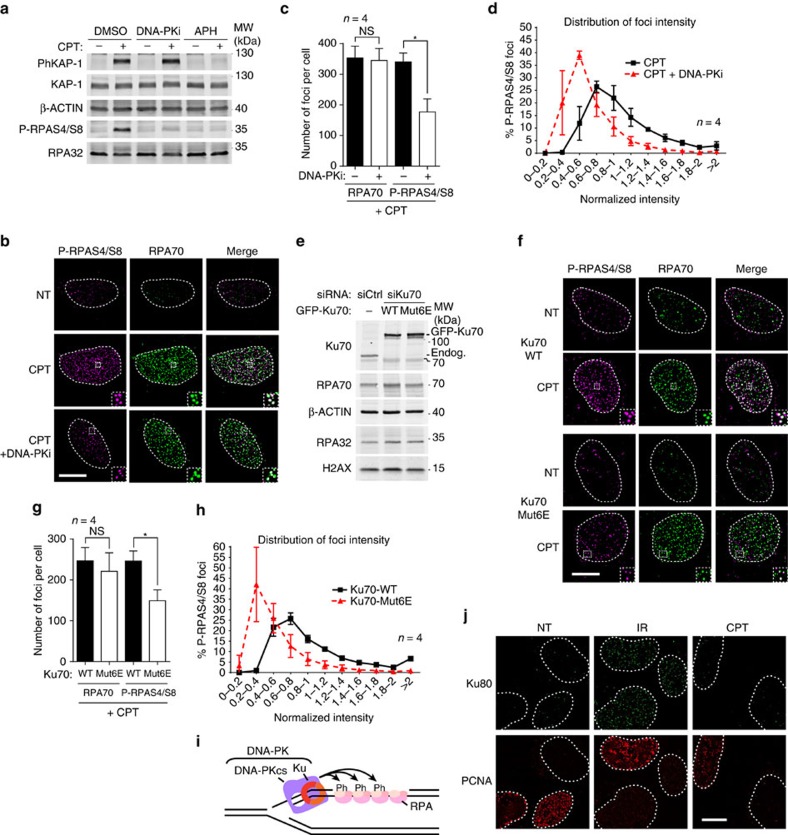
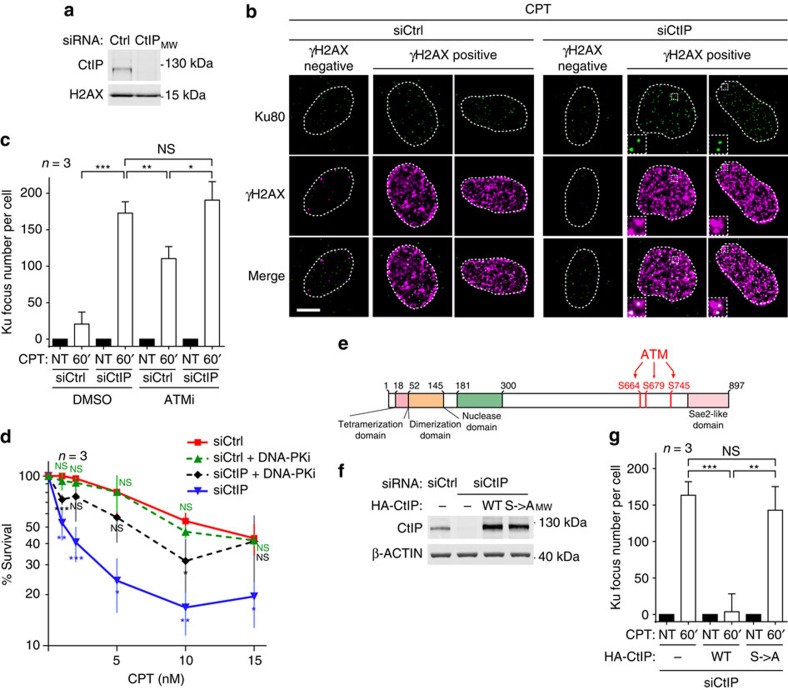
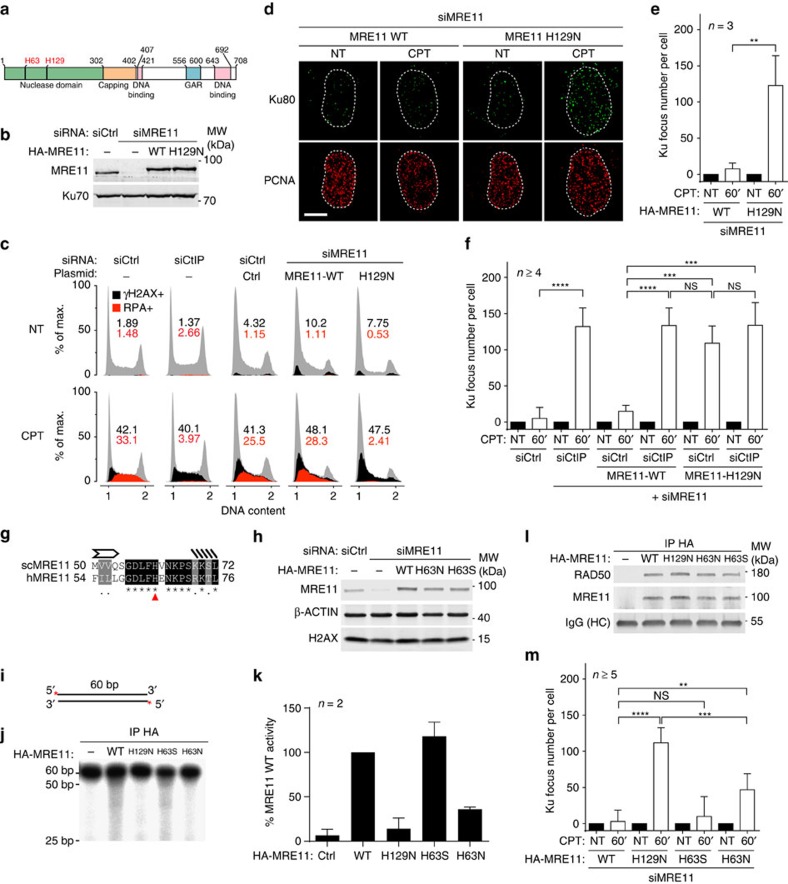
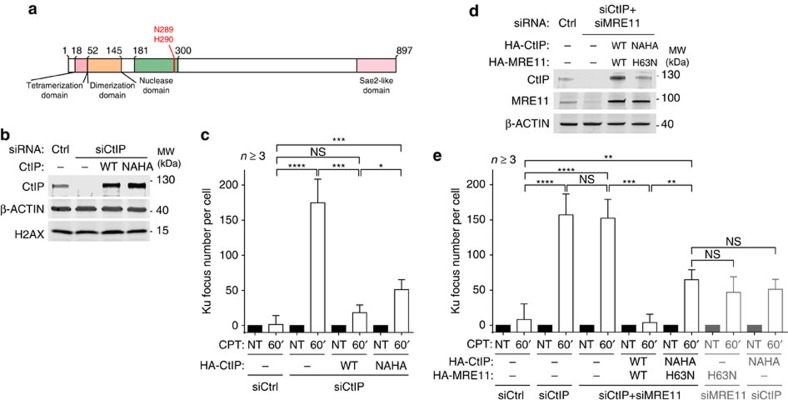
Repair of single-ended DNA double-strand breaks (seDSBs) by homologous recombination (HR) requires the generation of a 3' single-strand DNA overhang by exonuclease activities in a process called DNA resection. However, it is anticipated that the highly abundant DNA end-binding protein Ku sequesters seDSBs and shields them from exonuclease activities. Despite pioneering works in yeast, it is unclear how mammalian cells counteract Ku at seDSBs to allow HR to proceed. Here we show that in human cells, ATM-dependent phosphorylation of CtIP and the epistatic and coordinated actions of MRE11 and CtIP nuclease activities are required to limit the stable loading of Ku on seDSBs. We also provide evidence for a hitherto unsuspected additional mechanism that contributes to prevent Ku accumulation at seDSBs, acting downstream of MRE11 endonuclease activity and in parallel with MRE11 exonuclease activity. Finally, we show that Ku persistence at seDSBs compromises Rad51 focus assembly but not DNA resection.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
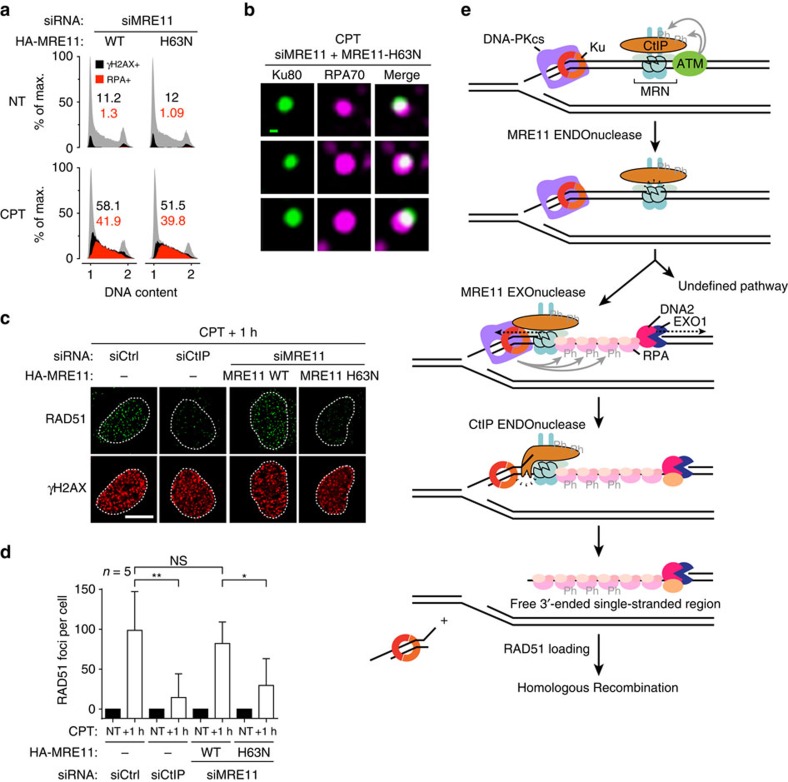
References
-
- Walker J. R., Corpina R. A. & Goldberg J. Structure of the Ku heterodimer bound to DNA and its implications for double-strand break repair. Nature 412, 607–614 (2001). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
