

Bmp15 Is an Oocyte-Produced Signal Required for Maintenance of the Adult Female Sexual Phenotype in Zebrafish
- PMID: 27642754
- PMCID: PMC5028036
- DOI: 10.1371/journal.pgen.1006323
Bmp15 Is an Oocyte-Produced Signal Required for Maintenance of the Adult Female Sexual Phenotype in Zebrafish
Abstract
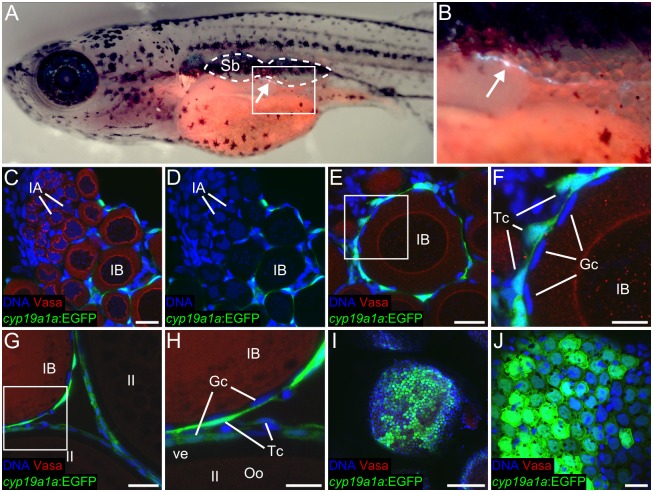
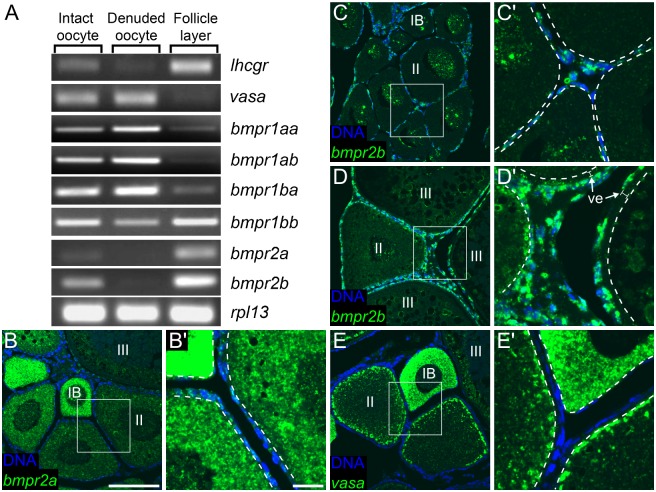
Although the zebrafish is a major model organism, how they determine sex is not well understood. In domesticated zebrafish, sex determination appears to be polygenic, being influenced by multiple genetic factors that may vary from strain to strain, and additionally can be influenced by environmental factors. However, the requirement of germ cells for female sex determination is well documented: animals that lack germ cells, or oocytes in particular, develop exclusively as males. Recently, it has been determined that oocytes are also required throughout the adult life of the animal to maintain the differentiated female state. How oocytes control sex differentiation and maintenance of the sexual phenotype is unknown. We therefore generated targeted mutations in genes for two oocyte produced signaling molecules, Bmp15 and Gdf9 and here report a novel role for Bmp15 in maintaining adult female sex differentiation in zebrafish. Females deficient in Bmp15 begin development normally but switch sex during the mid- to late- juvenile stage, and become fertile males. Additionally, by generating mutations in the aromatase cyp19a1a, we show that estrogen production is necessary for female development and that the function of Bmp15 in female sex maintenance is likely linked to the regulation of estrogen biosynthesis via promoting the development of estrogen-producing granulosa cells in the oocyte follicle.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




Similar articles
-
Cumulin, an Oocyte-secreted Heterodimer of the Transforming Growth Factor-β Family, Is a Potent Activator of Granulosa Cells and Improves Oocyte Quality.J Biol Chem. 2015 Sep 25;290(39):24007-20. doi: 10.1074/jbc.M115.671487. Epub 2015 Aug 8. J Biol Chem. 2015. PMID: 26254468 Free PMC article.
-
A variant of human growth differentiation factor-9 that improves oocyte developmental competence.J Biol Chem. 2020 Jun 5;295(23):7981-7991. doi: 10.1074/jbc.RA120.013050. Epub 2020 Apr 29. J Biol Chem. 2020. PMID: 32350111 Free PMC article.
-
Oocyte-Secreted Factors Synergize With FSH to Promote Aromatase Expression in Primary Human Cumulus Cells.J Clin Endocrinol Metab. 2019 May 1;104(5):1667-1676. doi: 10.1210/jc.2018-01705. J Clin Endocrinol Metab. 2019. PMID: 30541132 Free PMC article.
-
The role of oocyte-secreted factors GDF9 and BMP15 in follicular development and oogenesis.Reprod Domest Anim. 2011 Apr;46(2):354-61. doi: 10.1111/j.1439-0531.2010.01739.x. Epub 2010 Dec 30. Reprod Domest Anim. 2011. PMID: 21198974 Review.
-
Genetic regulation of sex determination and maintenance in zebrafish (Danio rerio).Curr Top Dev Biol. 2019;134:119-149. doi: 10.1016/bs.ctdb.2019.02.004. Epub 2019 Mar 21. Curr Top Dev Biol. 2019. PMID: 30999973 Free PMC article. Review.
Cited by
-
Shedding new light on early sex determination in zebrafish.Arch Toxicol. 2020 Dec;94(12):4143-4158. doi: 10.1007/s00204-020-02915-y. Epub 2020 Sep 25. Arch Toxicol. 2020. PMID: 32975586 Free PMC article.
-
The telomere bouquet is a hub where meiotic double-strand breaks, synapsis, and stable homolog juxtaposition are coordinated in the zebrafish, Danio rerio.PLoS Genet. 2019 Jan 17;15(1):e1007730. doi: 10.1371/journal.pgen.1007730. eCollection 2019 Jan. PLoS Genet. 2019. PMID: 30653507 Free PMC article.
-
Zebrafish as a Model for Germ Cell Regeneration.Front Cell Dev Biol. 2021 Jul 22;9:685001. doi: 10.3389/fcell.2021.685001. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34368134 Free PMC article. Review.
-
In Silico Prediction of Transcription Factor Collaborations Underlying Phenotypic Sexual Dimorphism in Zebrafish (Danio rerio).Genes (Basel). 2021 Jun 7;12(6):873. doi: 10.3390/genes12060873. Genes (Basel). 2021. PMID: 34200177 Free PMC article.
-
Microglia and Chek2 contribute to sex-specific organization of the adult zebrafish brain.bioRxiv [Preprint]. 2025 Aug 21:2025.08.15.670359. doi: 10.1101/2025.08.15.670359. bioRxiv. 2025. PMID: 40894757 Free PMC article. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
