

Roles of eukaryotic topoisomerases in transcription, replication and genomic stability
- PMID: 27649880
- PMCID: PMC9248348
- DOI: 10.1038/nrm.2016.111
Roles of eukaryotic topoisomerases in transcription, replication and genomic stability
Abstract
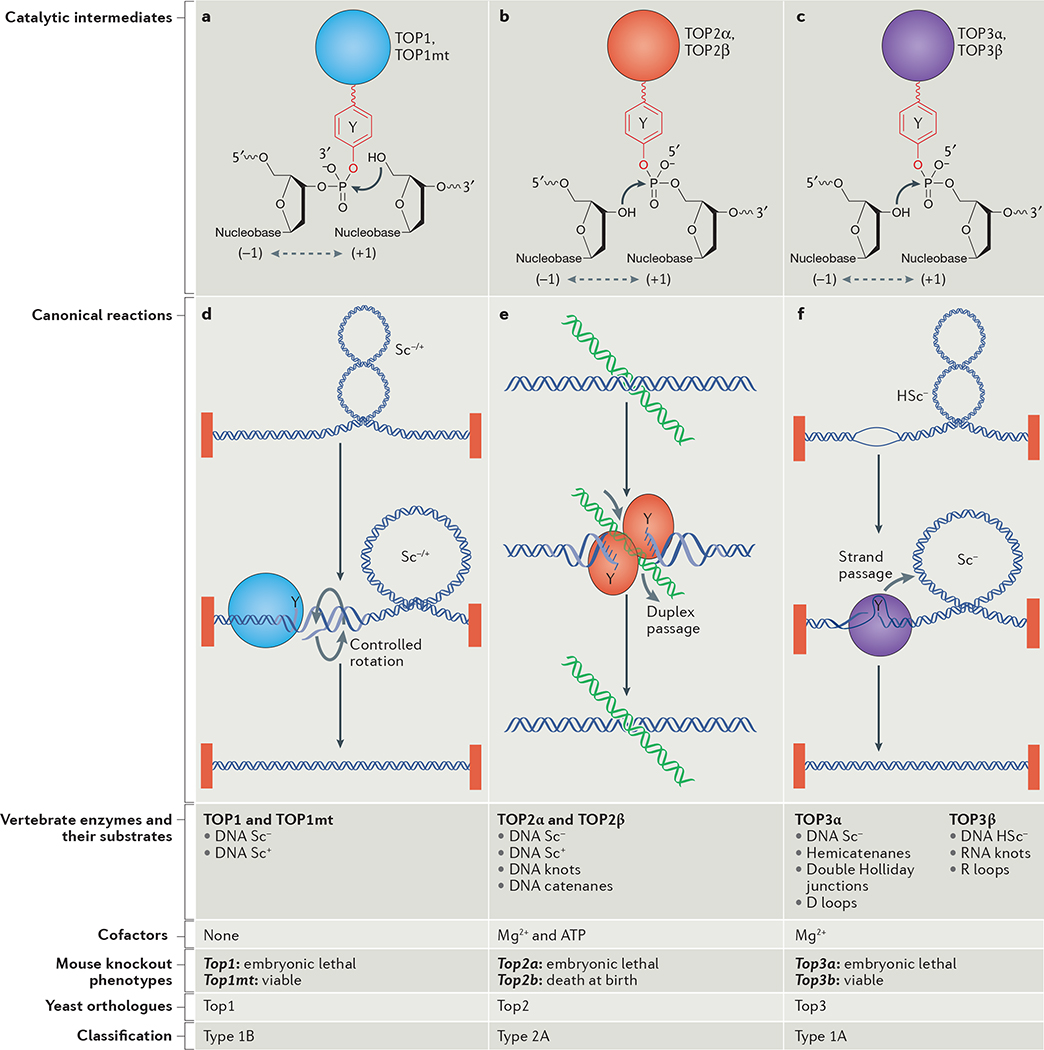
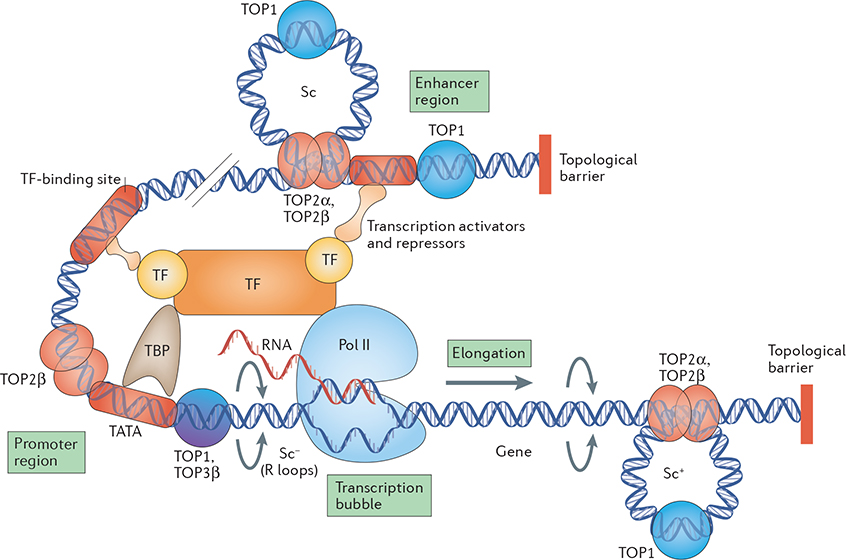
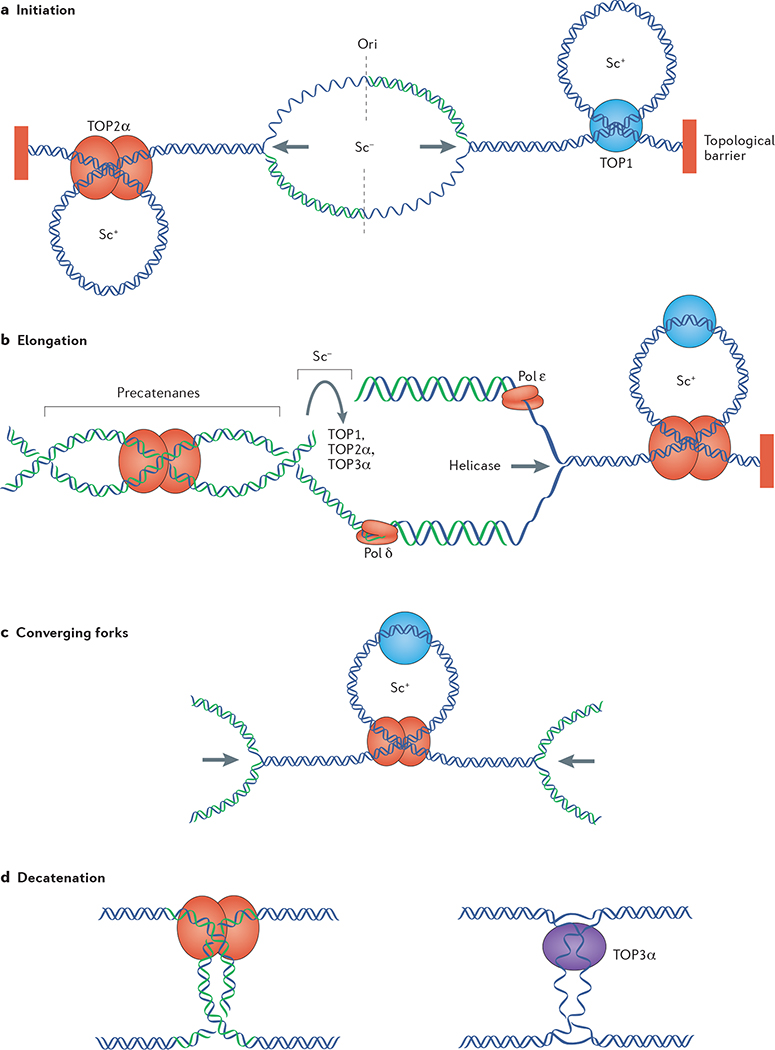
Topoisomerases introduce transient DNA breaks to relax supercoiled DNA, remove catenanes and enable chromosome segregation. Human cells encode six topoisomerases (TOP1, TOP1mt, TOP2α, TOP2β, TOP3α and TOP3β), which act on a broad range of DNA and RNA substrates at the nuclear and mitochondrial genomes. Their catalytic intermediates, the topoisomerase cleavage complexes (TOPcc), are therapeutic targets of various anticancer drugs. TOPcc can also form on damaged DNA during replication and transcription, and engage specific repair pathways, such as those mediated by tyrosyl-DNA phosphodiesterase 1 (TDP1) and TDP2 and by endonucleases (MRE11, XPF-ERCC1 and MUS81). Here, we review the roles of topoisomerases in mediating chromatin dynamics, transcription, replication, DNA damage repair and genomic stability, and discuss how deregulation of topoisomerases can cause neurodegenerative diseases, immune disorders and cancer.
Conflict of interest statement
Competing interests statement
The authors declare no competing interests.
Figures


References
-
- Watson JD & Crick FH Genetical implications of the structure of deoxyribonucleic acid. Nature 171, 964–967 (1953). - PubMed
-
- Wang JC Cellular roles of DNA topoisomerases: a molecular perspective. Nat. Rev. Mol. Cell Biol. 3, 430–440 (2002). - PubMed
-
- Chen SH, Chan NL & Hsieh TS New mechanistic and functional insights into DNA topoisomerases. Annu. Rev. Biochem. 82, 139–170 (2013). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
