

Polyamines release the let-7b-mediated suppression of initiation codon recognition during the protein synthesis of EXT2
- PMID: 27650265
- PMCID: PMC5030709
- DOI: 10.1038/srep33549
Polyamines release the let-7b-mediated suppression of initiation codon recognition during the protein synthesis of EXT2
Abstract
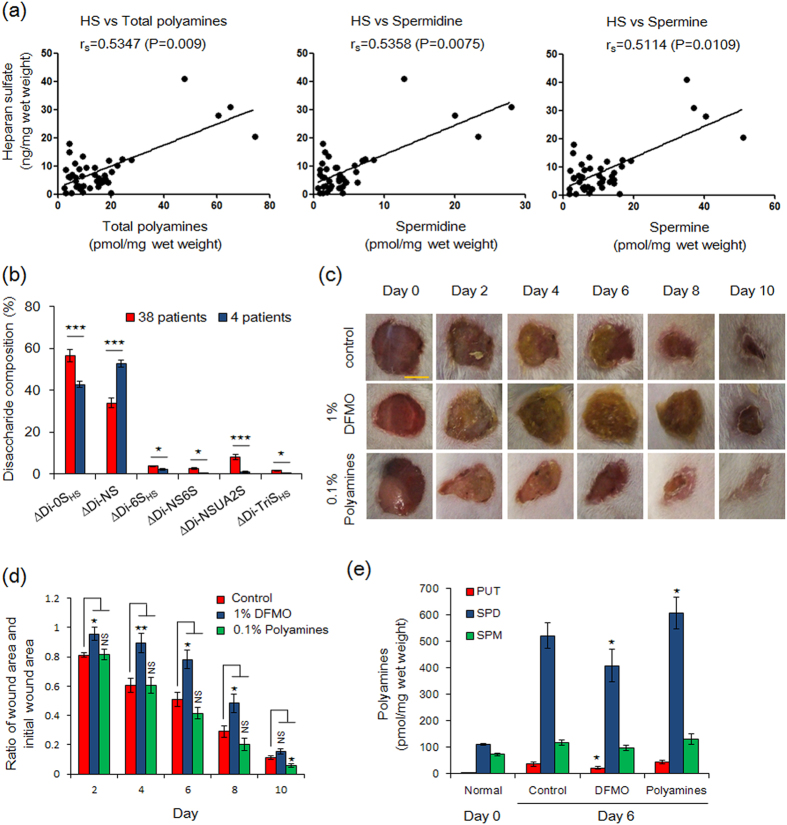
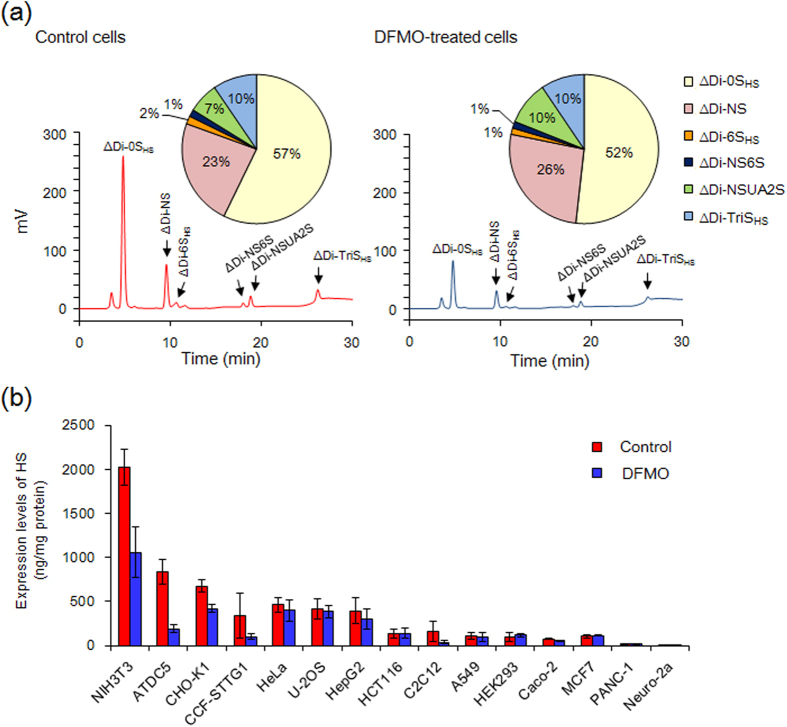
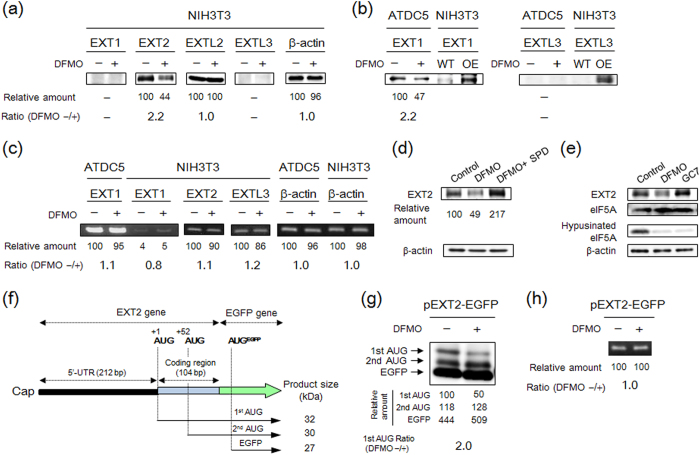
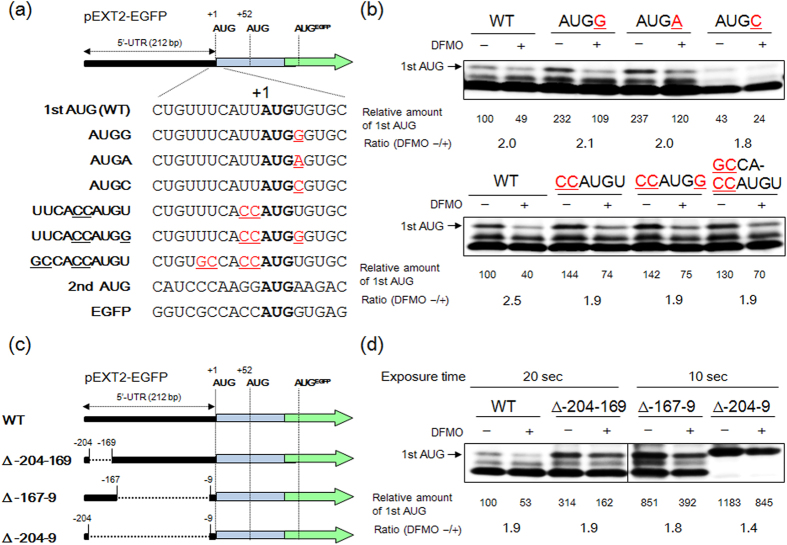
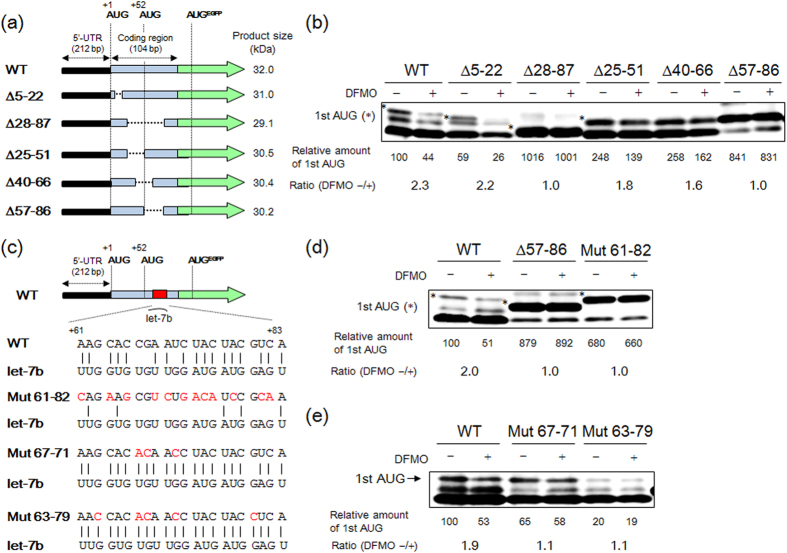
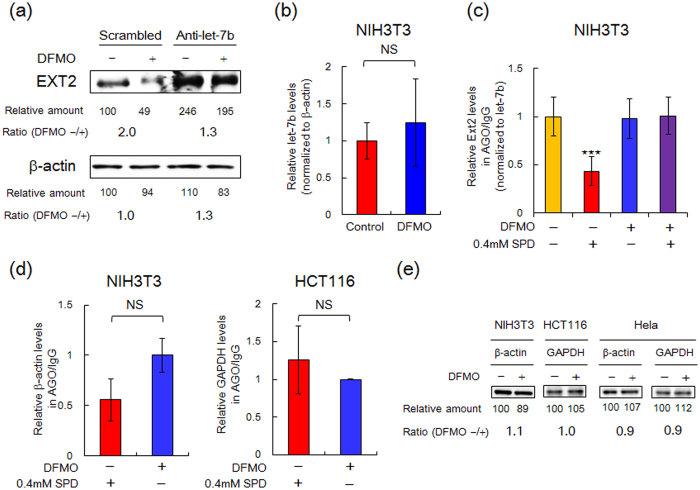
Proteoglycans (PGs), a family of glycosaminoglycan (GAG)-protein glycoconjugates, contribute to animal physiology through interactions between their glycan chains and growth factors, chemokines and adhesion molecules. However, it remains unclear how GAG structures are changed during the aging process. Here, we found that polyamine levels are correlated with the expression level of heparan sulfate (HS) in human skin. In cultured cell lines, the EXT1 and EXT2 enzymes, initiating HS biosynthesis, were stimulated at the translational level by polyamines. Interestingly, the initiation codon recognition by 43S preinitiation complex during EXT2 translation is suppressed by let-7b, a member of the let-7 microRNA family, through binding at the N-terminal amino acid coding sequence in EXT2 mRNA. Let-7b-mediated suppression of initiation codon depends on the length of 5'-UTR of EXT2 mRNA and its suppression is inhibited in the presence of polyamines. These findings provide new insights into the HS biosynthesis related to miRNA and polyamines.
Figures
References
-
- Roden L. Structure and metabolism of connective tissue proteoglycans. 267–371 (Plenum press, New York, 1980).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
