

BPTF Depletion Enhances T-cell-Mediated Antitumor Immunity
- PMID: 27651309
- PMCID: PMC5093041
- DOI: 10.1158/0008-5472.CAN-15-3125
BPTF Depletion Enhances T-cell-Mediated Antitumor Immunity
Abstract
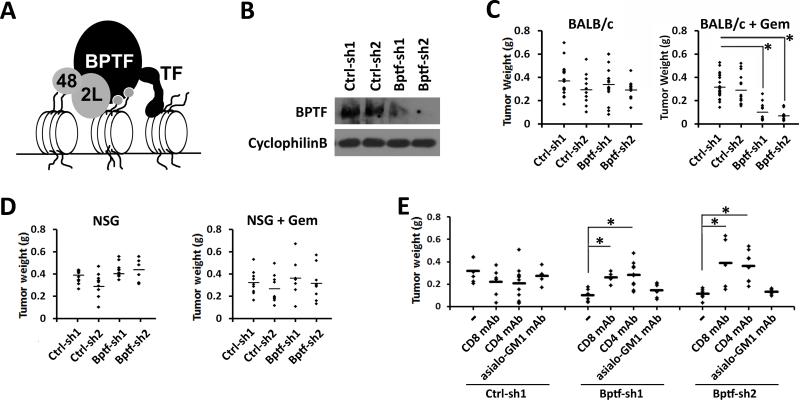
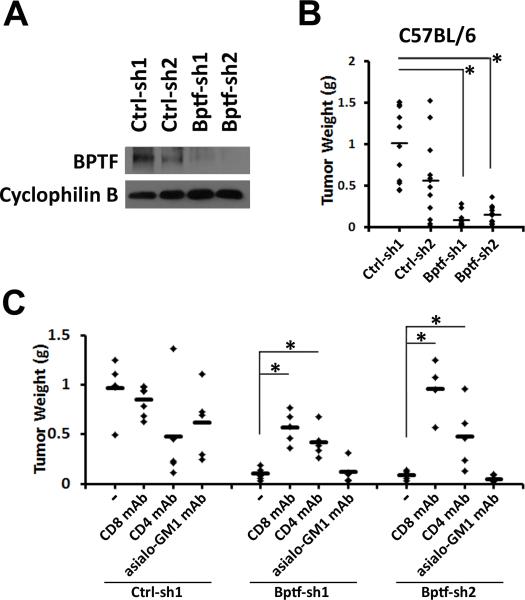
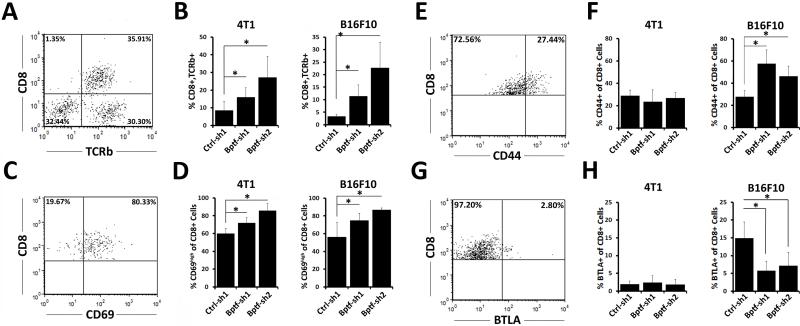
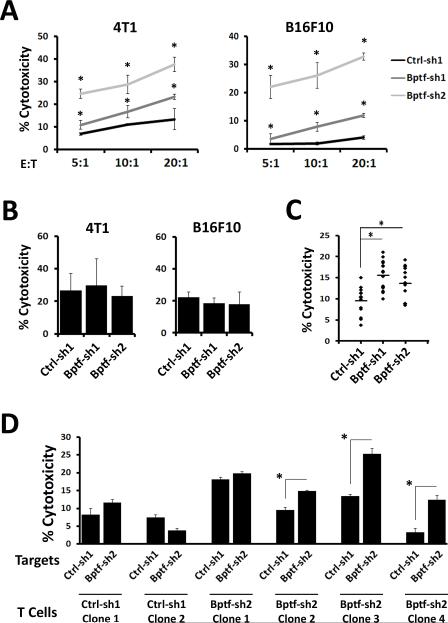
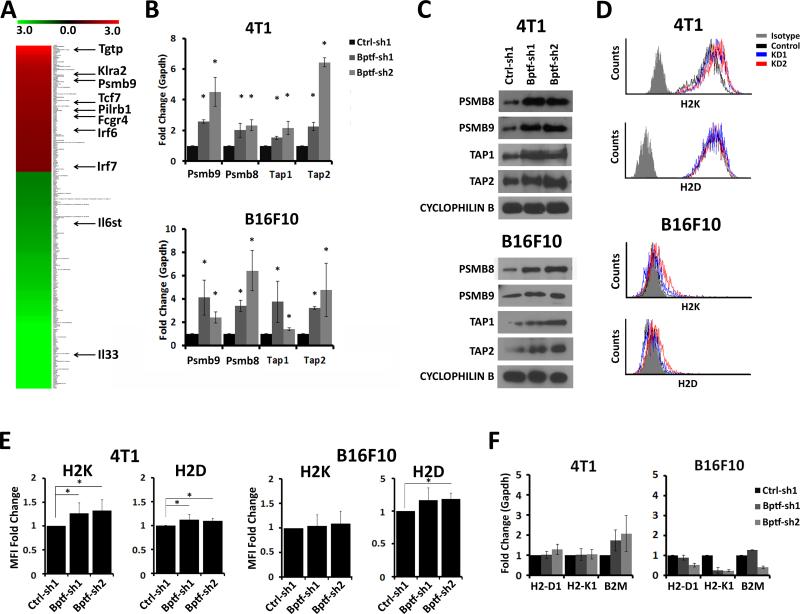
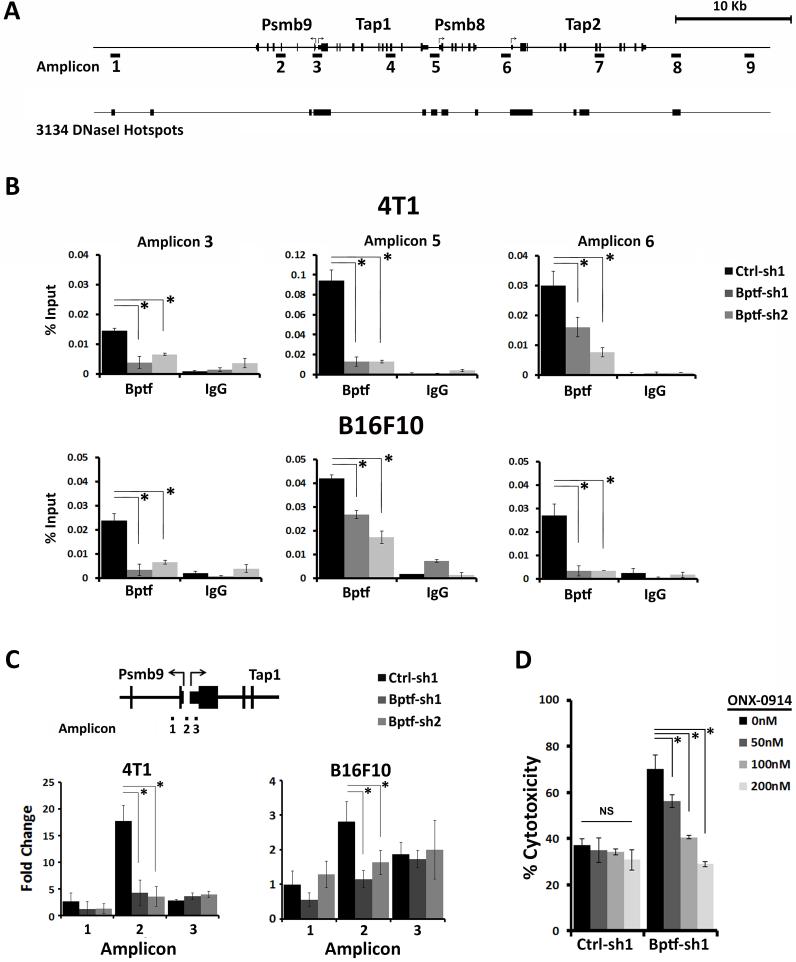
Genetic studies in fruit flies have implicated the chromatin remodeling complex nucleosome remodeling factor (NURF) in immunity, but it has yet to be studied in mammals. Here we show that its targeting in mice enhances antitumor immunity in two syngeneic models of cancer. NURF was disabled by silencing of bromodomain PHD-finger containing transcription factor (BPTF), the largest and essential subunit of NURF. We found that both CD8+ and CD4+ T cells were necessary for enhanced antitumor activity, with elevated numbers of activated CD8+ T cells observed in BPTF-deficient tumors. Enhanced cytolytic activity was observed for CD8+ T cells cocultured with BPTF-silenced cells. Similar effects were not produced with T-cell receptor transgenic CD8+ T cells, implicating the involvement of novel antigens. Accordingly, enhanced activity was observed for individual CD8+ T-cell clones from mice bearing BPTF-silenced tumors. Mechanistic investigations revealed that NURF directly regulated the expression of genes encoding immunoproteasome subunits Psmb8 and Psmb9 and the antigen transporter genes Tap1 and Tap2 The PSMB8 inhibitor ONX-0914 reversed the effects of BPTF ablation, consistent with a critical role for the immunoproteasome in improving tumor immunogenicity. Thus, NURF normally suppresses tumor antigenicity and its depletion improves antigen processing, CD8 T-cell cytotoxicity, and antitumor immunity, identifying NURF as a candidate therapeutic target to enhance antitumor immunity. Cancer Res; 76(21); 6183-92. ©2016 AACR.
©2016 American Association for Cancer Research.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
