

Characteristic Response to Chemosensory Signals in GABAergic Cells of Medial Amygdala Is Not Driven by Main Olfactory Input
- PMID: 27651427
- PMCID: PMC5155562
- DOI: 10.1093/chemse/bjw096
Characteristic Response to Chemosensory Signals in GABAergic Cells of Medial Amygdala Is Not Driven by Main Olfactory Input
Abstract
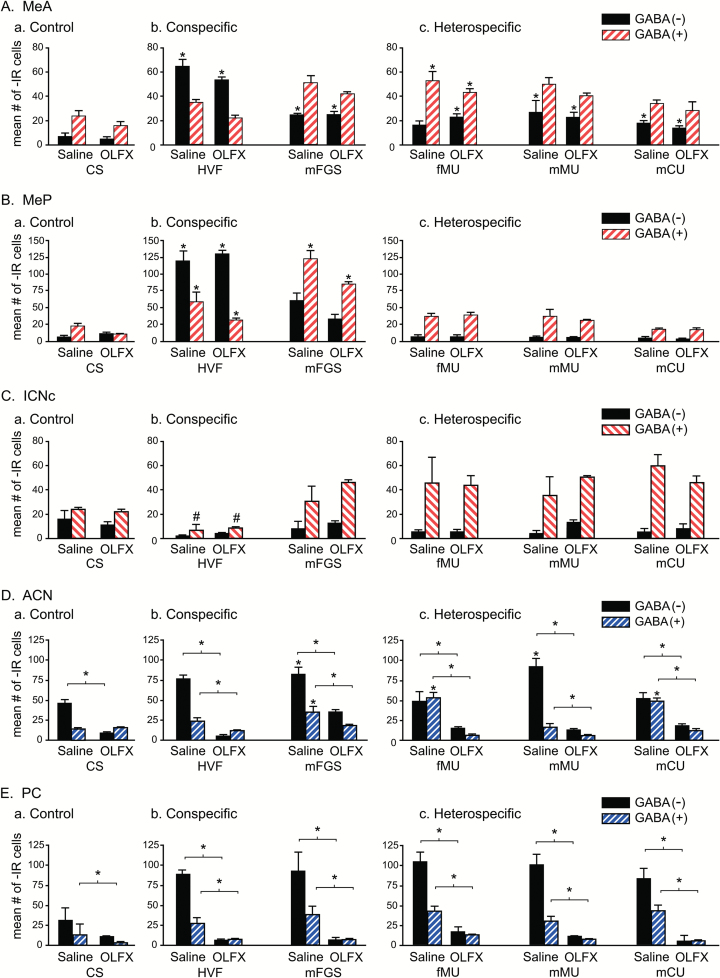
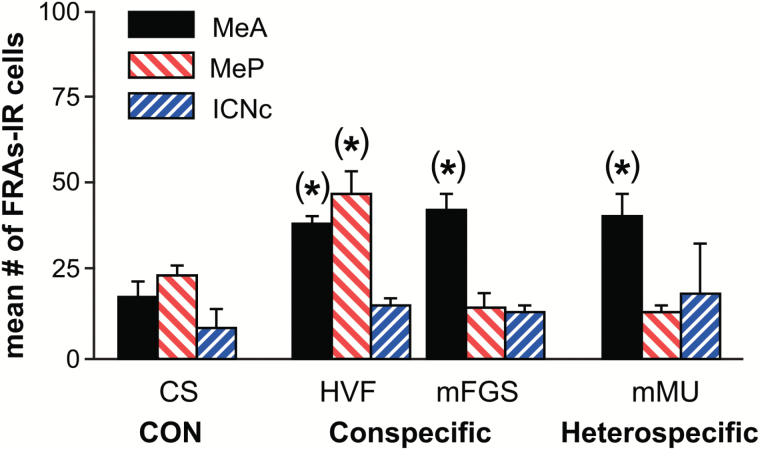
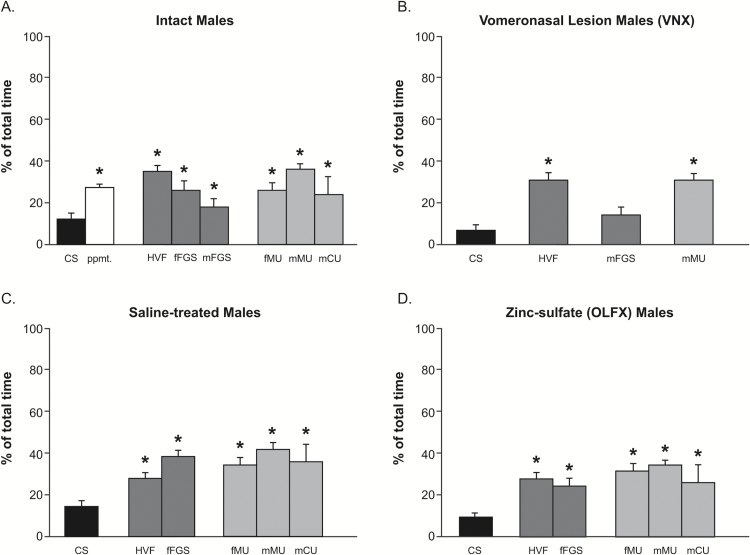
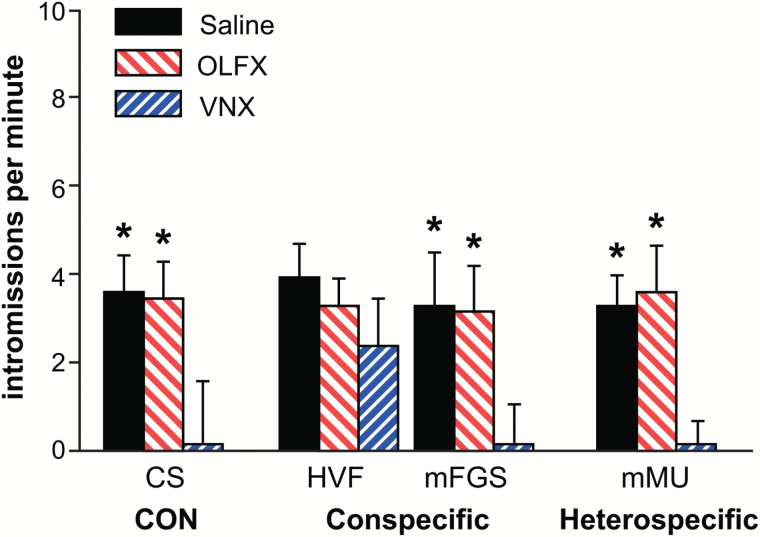
Chemosensory stimuli from same species (conspecific) and different species (heterospecific) elicit categorically different immediate-early gene (IEG) response patterns in medial amygdala in male hamsters and mice. All heterospecific stimuli activate anterior medial amygdala (MeA) but only especially salient heterospecific stimuli, such as those from predators activate posterior medial amygdala (MeP). We previously reported that characteristic patterns of response in separate populations of cells in MeA and MeP distinguish between different conspecific stimuli. Both gamma aminobutyric acid (GABA)-immunoreactive (ir) cells and GABA-receptor-ir cells make this distinction. Here, using zinc sulfate lesions of the main olfactory epithelium, we show evidence that main olfactory input does not contribute to the characteristic patterns of response in GABA-ir cells of male hamster amygdala, either for conspecific or heterospecific stimuli. Some GABAergic cells are output neurons carrying information from medial amygdala to behavioral executive regions of basal forebrain. Thus, the differential response to different conspecific signals can lead to differential activation of downstream circuits based on nonolfactory input. Finally, we show that an intact vomeronasal organ is necessary and sufficient to produce the characteristic patterns of response to conspecific and heterospecific chemosensory stimuli in hamster medial amygdala. Although main olfactory input may be critical in species with less prominent vomeronasal input for equivalent medial amygdala responses, work presented here suggests that hamster medial amygdala uses primarily vomeronasal input to discriminate between important unlearned conspecific social signals, to distinguish them from the social signals of other species, and may convey that information to brain circuits eliciting appropriate social behavior.
Keywords: GABA; IEG; chemosignal; circuit; lesion; vomeronasal.
© The Author 2016. Published by Oxford University Press. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures
References
-
- Baum MJ, Cherry JA. 2015. Processing by the main olfactory system of chemosignals that facilitate mammalian reproduction. Horm Behav. 68:53–64. - PubMed
-
- Baum MJ, Kelliher KR. 2009. Complementary roles of the main and accessory olfactory systems in mammalian mate recognition. Annu Rev Physiol. 71:141–160. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
