

Autonomous translational pausing is required for XBP1u mRNA recruitment to the ER via the SRP pathway
- PMID: 27651490
- PMCID: PMC5056097
- DOI: 10.1073/pnas.1604435113
Autonomous translational pausing is required for XBP1u mRNA recruitment to the ER via the SRP pathway
Abstract
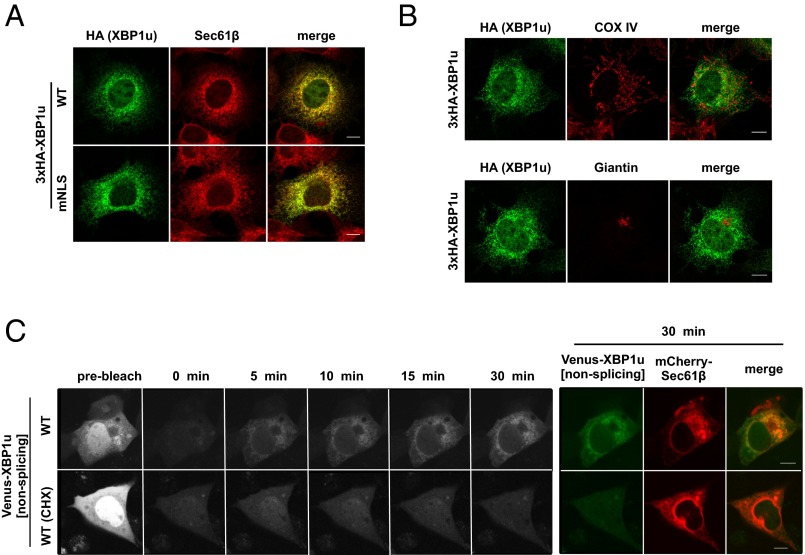
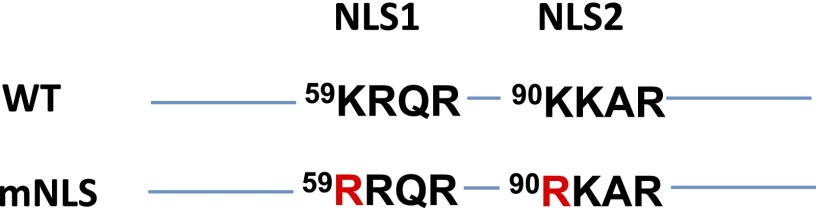
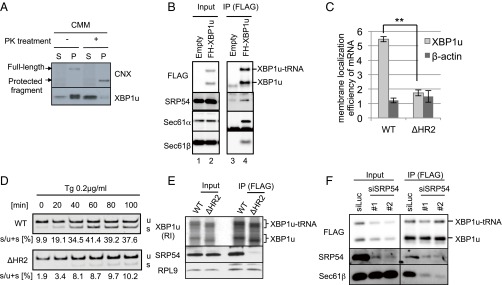
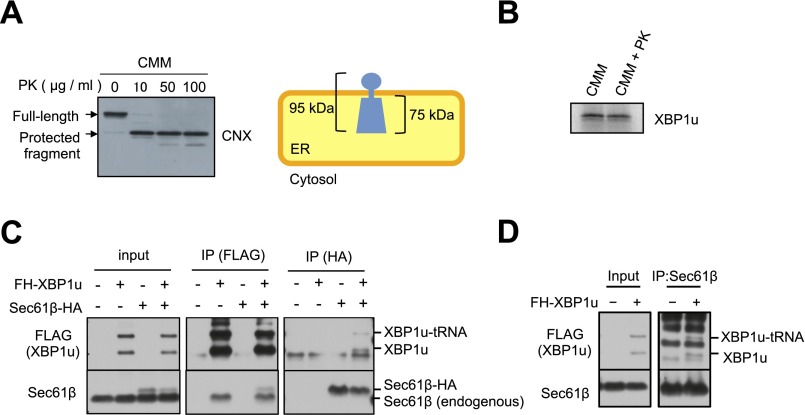
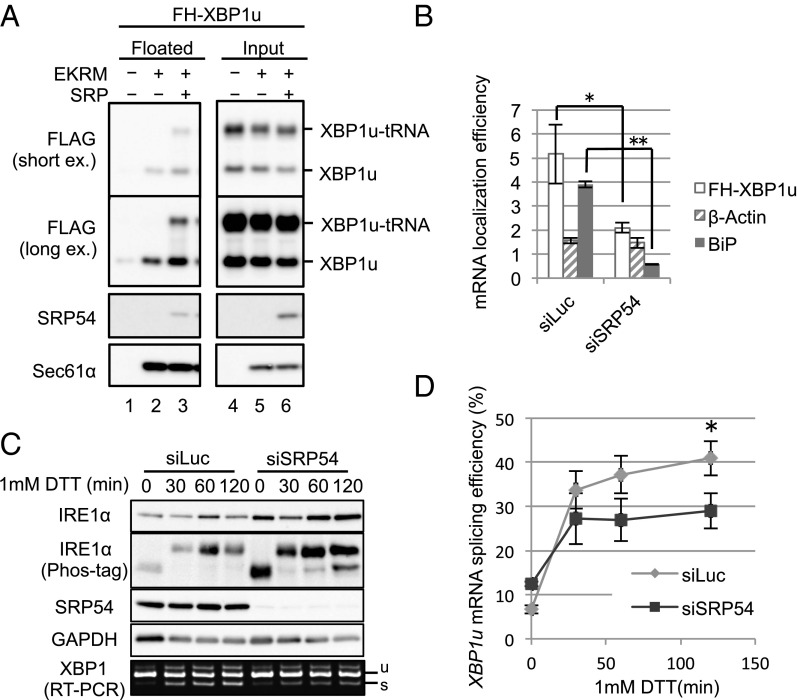
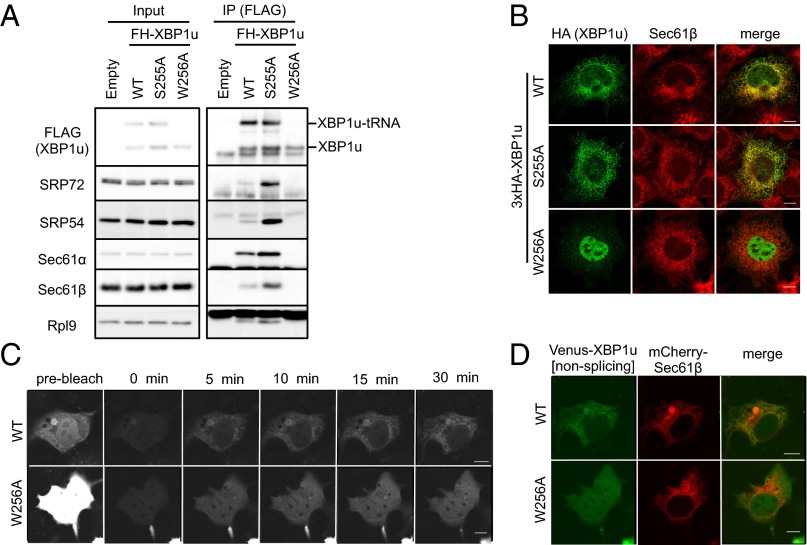
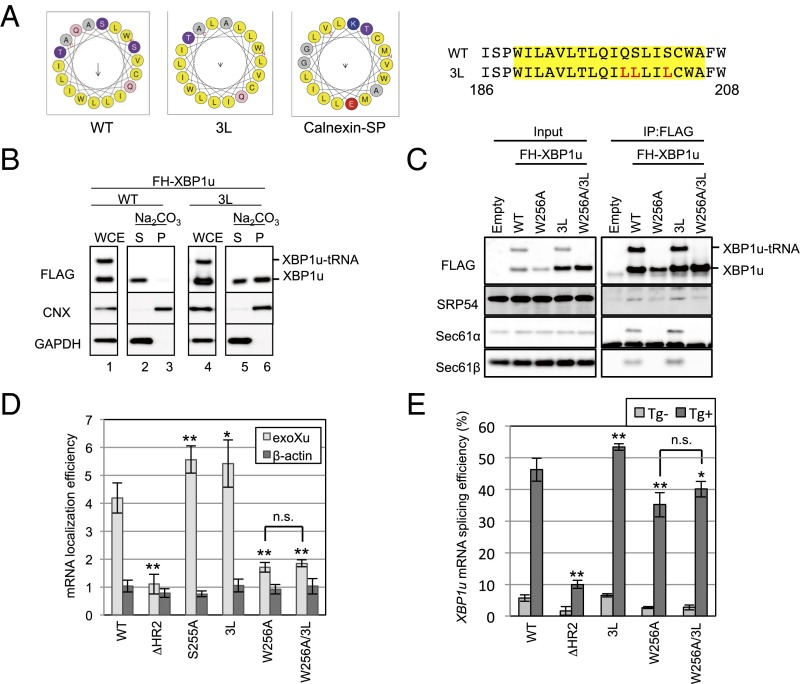
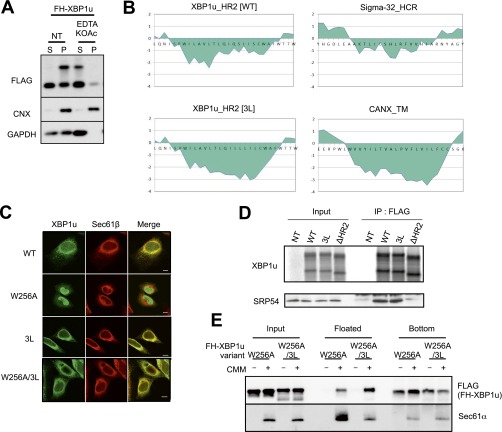
Unconventional mRNA splicing on the endoplasmic reticulum (ER) membrane is the sole conserved mechanism in eukaryotes to transmit information regarding misfolded protein accumulation to the nucleus to activate the stress response. In metazoans, the unspliced form of X-box-binding protein 1 (XBP1u) mRNA is recruited to membranes as a ribosome nascent chain (RNC) complex for efficient splicing. We previously reported that both hydrophobic (HR2) and translational pausing regions of XBP1u are important for the recruitment of its own mRNA to membranes. However, its precise location and the molecular mechanism of translocation are unclear. We show that XBP1u-RNC is specifically recruited to the ER membrane in an HR2- and translational pausing-dependent manner by immunostaining, fluorescent recovery after photobleaching, and biochemical analyses. Notably, translational pausing during XBP1u synthesis is indispensable for the recognition of HR2 by the signal recognition particle (SRP), resulting in efficient ER-specific targeting of the complex, similar to secretory protein targeting to the ER. On the ER, the XBP1u nascent chain is transferred from the SRP to the translocon; however, it cannot pass through the translocon or insert into the membrane. Therefore, our results support a noncanonical mechanism by which mRNA substrates are recruited to the ER for unconventional splicing.
Keywords: SRP; XBP1 mRNA; translational pausing; translocon; unfolded protein response.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Translational pausing ensures membrane targeting and cytoplasmic splicing of XBP1u mRNA.Science. 2011 Feb 4;331(6017):586-9. doi: 10.1126/science.1197142. Epub 2011 Jan 13. Science. 2011. PMID: 21233347
-
Structural and mutational analysis of the ribosome-arresting human XBP1u.Elife. 2019 Jun 27;8:e46267. doi: 10.7554/eLife.46267. Elife. 2019. PMID: 31246176 Free PMC article.
-
Cotranslational targeting of XBP1 protein to the membrane promotes cytoplasmic splicing of its own mRNA.Mol Cell. 2009 Apr 24;34(2):191-200. doi: 10.1016/j.molcel.2009.02.033. Mol Cell. 2009. PMID: 19394296
-
Co-translational targeting and translocation of proteins to the endoplasmic reticulum.Biochim Biophys Acta. 2013 Nov;1833(11):2392-402. doi: 10.1016/j.bbamcr.2013.02.021. Epub 2013 Feb 26. Biochim Biophys Acta. 2013. PMID: 23481039 Review.
-
Structure, function and evolution of the signal recognition particle.EMBO J. 2003 Jul 15;22(14):3479-85. doi: 10.1093/emboj/cdg337. EMBO J. 2003. PMID: 12853463 Free PMC article. Review.
Cited by
-
Distinct Functions of Acyl/Alkyl Dihydroxyacetonephosphate Reductase in Peroxisomes and Endoplasmic Reticulum.Front Cell Dev Biol. 2020 Sep 11;8:855. doi: 10.3389/fcell.2020.00855. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 33042986 Free PMC article.
-
Exploring the IRE1 interactome: From canonical signaling functions to unexpected roles.J Biol Chem. 2024 Apr;300(4):107169. doi: 10.1016/j.jbc.2024.107169. Epub 2024 Mar 15. J Biol Chem. 2024. PMID: 38494075 Free PMC article. Review.
-
Hepatocyte-specific GDF15 overexpression improves high-fat diet-induced obesity and hepatic steatosis in mice via hepatic FGF21 induction.Sci Rep. 2024 Oct 14;14(1):23993. doi: 10.1038/s41598-024-75107-8. Sci Rep. 2024. PMID: 39402176 Free PMC article.
-
Phylogeny-linked occurrence of ribosome stalling on the mRNAs of Arabidopsis unfolded protein response factor bZIP60 orthologs in divergent plant species.Nucleic Acids Res. 2024 May 8;52(8):4276-4294. doi: 10.1093/nar/gkae101. Nucleic Acids Res. 2024. PMID: 38366760 Free PMC article.
-
HTS Identification of Activators and Inhibitors of Endoplasmic Reticulum (ER) Stress and the Unfolded Protein Response (UPR).Methods Mol Biol. 2022;2378:317-327. doi: 10.1007/978-1-0716-1732-8_20. Methods Mol Biol. 2022. PMID: 34985709
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
