

The Effect of Temperature on Microtubule-Based Transport by Cytoplasmic Dynein and Kinesin-1 Motors
- PMID: 27653487
- PMCID: PMC5034348
- DOI: 10.1016/j.bpj.2016.08.006
The Effect of Temperature on Microtubule-Based Transport by Cytoplasmic Dynein and Kinesin-1 Motors
Erratum in
-
The Effect of Temperature on Microtubule-Based Transport by Cytoplasmic Dynein and Kinesin-1 Motors.Biophys J. 2016 Oct 18;111(8):1816. doi: 10.1016/j.bpj.2016.09.040. Biophys J. 2016. PMID: 27760367 Free PMC article. No abstract available.
Abstract
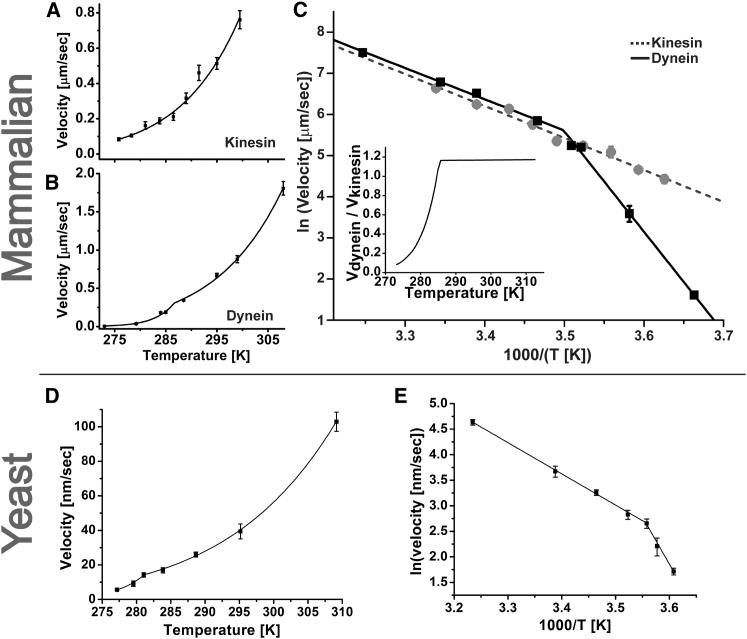
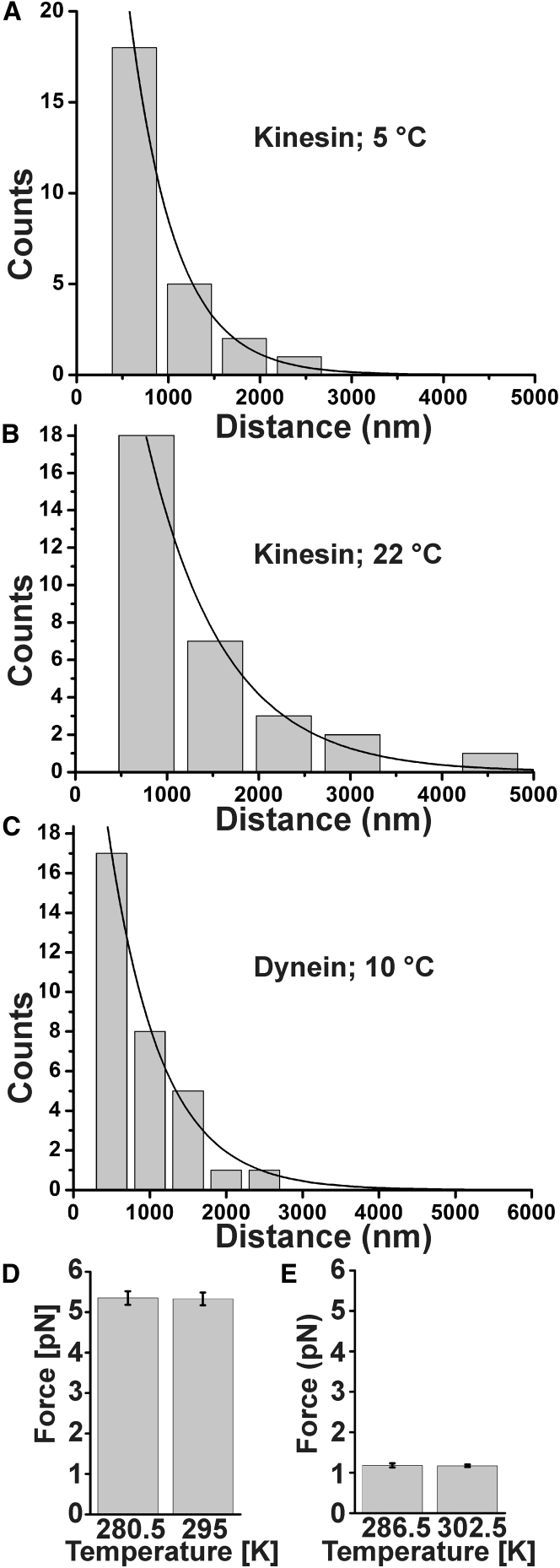
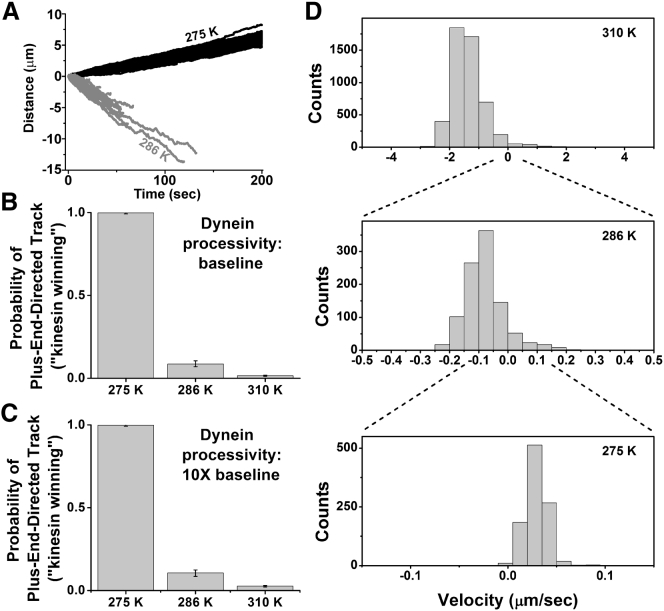
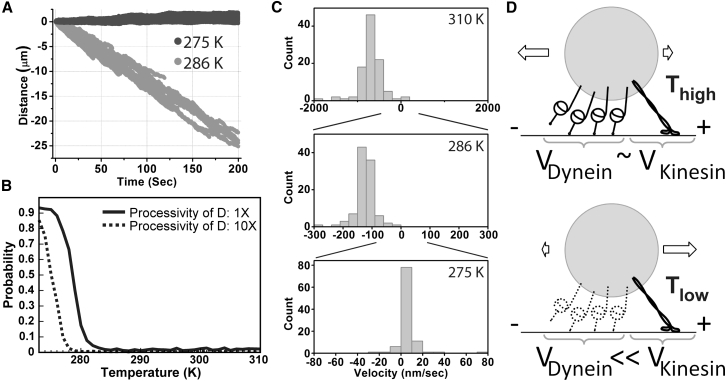
Cytoplasmic dynein and kinesin are both microtubule-based molecular motors but are structurally and evolutionarily unrelated. Under standard conditions, both move with comparable unloaded velocities toward either the microtubule minus (dynein) or plus (most kinesins) end. This similarity is important because it is often implicitly incorporated into models that examine the balance of cargo fluxes in cells and into models of the bidirectional motility of individual cargos. We examined whether this similarity is a robust feature, and specifically whether it persists across the biologically relevant temperature range. The velocity of mammalian cytoplasmic dynein, but not of mammalian kinesin-1, exhibited a break from simple Arrhenius behavior below 15°C-just above the restrictive temperature of mammalian fast axonal transport. In contrast, the velocity of yeast cytoplasmic dynein showed a break from Arrhenius behavior at a lower temperature (∼8°C). Our studies implicate cytoplasmic dynein as a more thermally tunable motor and therefore a potential thermal regulator of microtubule-based transport. Our theoretical analysis further suggests that motor velocity changes can lead to qualitative changes in individual cargo motion and hence net intracellular cargo fluxes. We propose that temperature can potentially be used as a noninvasive probe of intracellular transport.
Copyright © 2016 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Carey H.V., Andrews M.T., Martin S.L. Mammalian hibernation: cellular and molecular responses to depressed metabolism and low temperature. Physiol. Rev. 2003;83:1153–1181. - PubMed
-
- Cancalon P. Influence of temperature on various mechanisms associated with neuronal growth and nerve regeneration. Prog. Neurobiol. 1985;25:27–92. - PubMed
-
- Mallik R., Rai A.K., Kunwar A. Teamwork in microtubule motors. Trends Cell Biol. 2013;23:575–582. - PubMed
-
- Gross S.P., Vershinin M., Shubeita G.T. Cargo transport: two motors are sometimes better than one. Curr. Biol. 2007;17:R478–R486. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
