

Bcl11a Deficiency Leads to Hematopoietic Stem Cell Defects with an Aging-like Phenotype
- PMID: 27653684
- PMCID: PMC5054719
- DOI: 10.1016/j.celrep.2016.08.064
Bcl11a Deficiency Leads to Hematopoietic Stem Cell Defects with an Aging-like Phenotype
Abstract
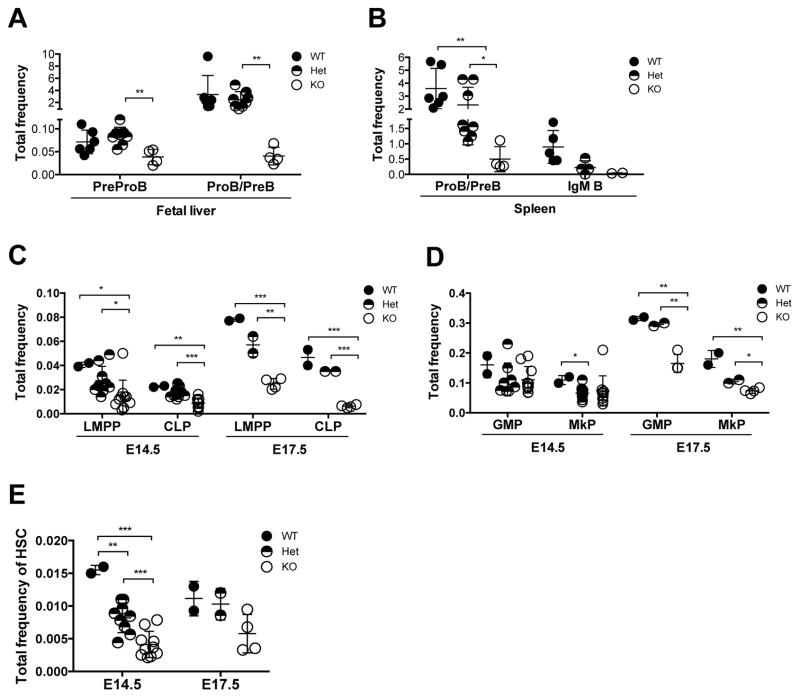
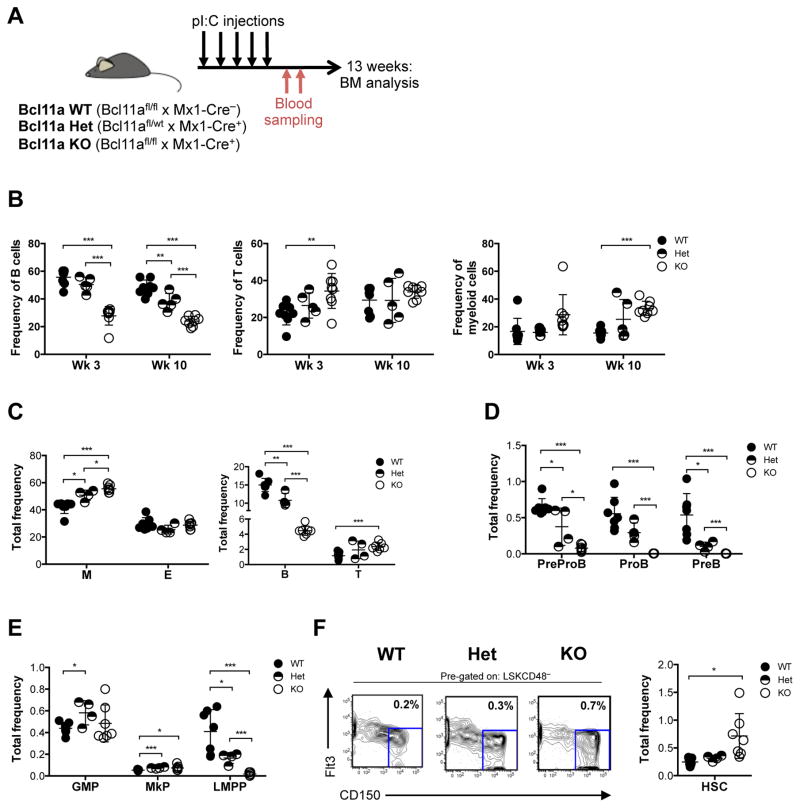
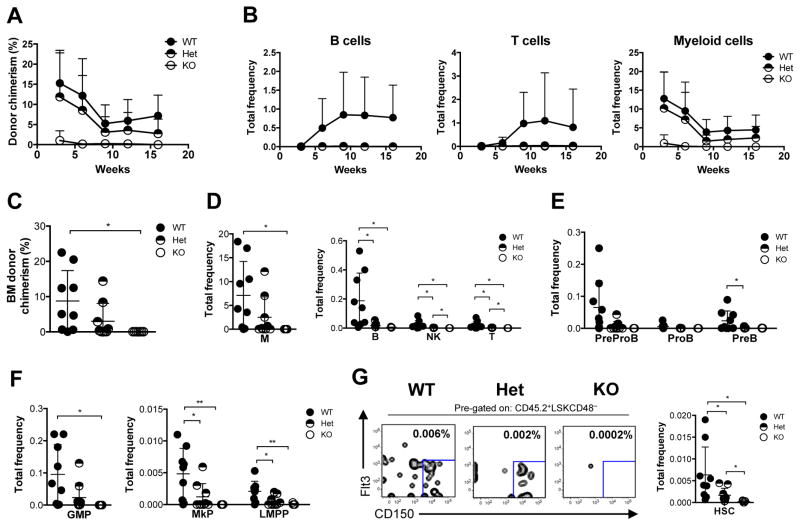
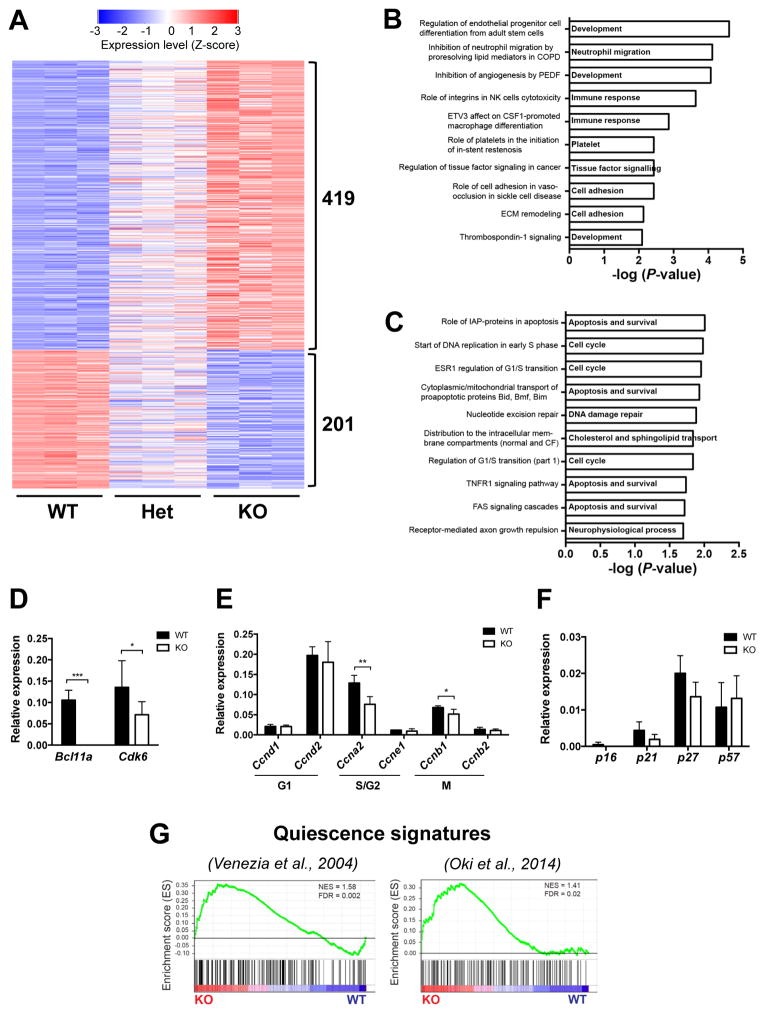
B cell CLL/lymphoma 11A (BCL11A) is a transcription factor and regulator of hemoglobin switching that has emerged as a promising therapeutic target for sickle cell disease and thalassemia. In the hematopoietic system, BCL11A is required for B lymphopoiesis, yet its role in other hematopoietic cells, especially hematopoietic stem cells (HSCs) remains elusive. The extensive expression of BCL11A in hematopoiesis implicates context-dependent roles, highlighting the importance of fully characterizing its function as part of ongoing efforts for stem cell therapy and regenerative medicine. Here, we demonstrate that BCL11A is indispensable for normal HSC function. Bcl11a deficiency results in HSC defects, typically observed in the aging hematopoietic system. We find that downregulation of cyclin-dependent kinase 6 (Cdk6), and the ensuing cell-cycle delay, correlate with HSC dysfunction. Our studies define a mechanism for BCL11A in regulation of HSC function and have important implications for the design of therapeutic approaches to targeting BCL11A.
Copyright © 2016 The Authors. Published by Elsevier Inc. All rights reserved.
Figures



Similar articles
-
Lineage-specific BCL11A knockdown circumvents toxicities and reverses sickle phenotype.J Clin Invest. 2016 Oct 3;126(10):3868-3878. doi: 10.1172/JCI87885. Epub 2016 Sep 6. J Clin Invest. 2016. PMID: 27599293 Free PMC article.
-
Single-cell transcriptomic reconstruction reveals cell cycle and multi-lineage differentiation defects in Bcl11a-deficient hematopoietic stem cells.Genome Biol. 2015 Sep 21;16:178. doi: 10.1186/s13059-015-0739-5. Genome Biol. 2015. PMID: 26387834 Free PMC article.
-
Bcl11a is essential for lymphoid development and negatively regulates p53.J Exp Med. 2012 Dec 17;209(13):2467-83. doi: 10.1084/jem.20121846. Epub 2012 Dec 10. J Exp Med. 2012. PMID: 23230003 Free PMC article.
-
Extracellular Vesicles and Hematopoietic Stem Cell Aging.Arterioscler Thromb Vasc Biol. 2021 Aug;41(8):e399-e416. doi: 10.1161/ATVBAHA.120.314643. Epub 2021 Jun 3. Arterioscler Thromb Vasc Biol. 2021. PMID: 34078091 Review.
-
Molecular mechanisms underlying lineage bias in aging hematopoiesis.Semin Hematol. 2017 Jan;54(1):4-11. doi: 10.1053/j.seminhematol.2016.11.002. Epub 2016 Nov 20. Semin Hematol. 2017. PMID: 28088987 Review.
Cited by
-
Strict in vivo specificity of the Bcl11a erythroid enhancer.Blood. 2016 Nov 10;128(19):2338-2342. doi: 10.1182/blood-2016-08-736249. Epub 2016 Oct 5. Blood. 2016. PMID: 27707736 Free PMC article.
-
Smarca5-mediated epigenetic programming facilitates fetal HSPC development in vertebrates.Blood. 2021 Jan 14;137(2):190-202. doi: 10.1182/blood.2020005219. Blood. 2021. PMID: 32756943 Free PMC article.
-
BCL11A enhances stemness and promotes progression by activating Wnt/β-catenin signaling in breast cancer.Cancer Manag Res. 2019 Apr 11;11:2997-3007. doi: 10.2147/CMAR.S199368. eCollection 2019. Cancer Manag Res. 2019. PMID: 31114347 Free PMC article.
-
Genome editing strategies for fetal hemoglobin induction in beta-hemoglobinopathies.Hum Mol Genet. 2020 Sep 30;29(R1):R100-R106. doi: 10.1093/hmg/ddaa088. Hum Mol Genet. 2020. PMID: 32406490 Free PMC article. Review.
-
Disruption of the BCL11A Erythroid Enhancer Reactivates Fetal Hemoglobin in Erythroid Cells of Patients with β-Thalassemia Major.Mol Ther Methods Clin Dev. 2018 Aug 14;10:313-326. doi: 10.1016/j.omtm.2018.08.003. eCollection 2018 Sep 21. Mol Ther Methods Clin Dev. 2018. PMID: 30182035 Free PMC article.
References
-
- Alcalay M, Meani N, Gelmetti V, Fantozzi A, Fagioli M, Orleth A, Riganelli D, Sebastiani C, Cappelli E, Casciari C, Sciurpi MT, Mariano AR, Minardi SP, Luzi L, Muller H, Di Fiore PP, Frosina G, Pelicci PG. Acute myeloid leukemia fusion proteins deregulate genes involved in stem cell maintenance and DNA repair. Journal of Clinical Investigation. 2003;112:1751–1761. doi: 10.1172/JCI200317595. - DOI - PMC - PubMed
-
- Bauer DE, Kamran SC, Lessard S, Xu J, Fujiwara Y, Lin C, Shao Z, Canver MC, Smith EC, Pinello L, Sabo PJ, Vierstra J, Voit RA, Yuan GC, Porteus MH, Stamatoyannopoulos JA, Lettre G, Orkin SH. An erythroid enhancer of BCL11A subject to genetic variation determines fetal hemoglobin level. Science. 2013;342:253–257. doi: 10.1126/science.1242088. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
