

Global Analysis of O-GlcNAc Glycoproteins in Activated Human T Cells
- PMID: 27655845
- PMCID: PMC5055199
- DOI: 10.4049/jimmunol.1502031
Global Analysis of O-GlcNAc Glycoproteins in Activated Human T Cells
Abstract
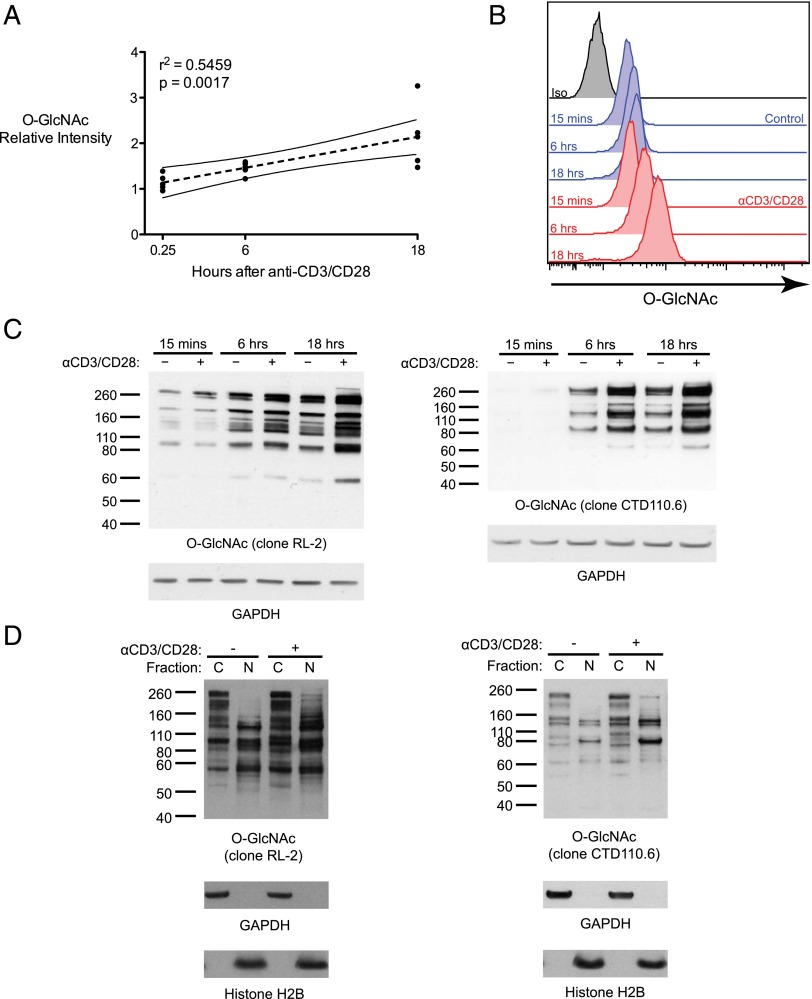
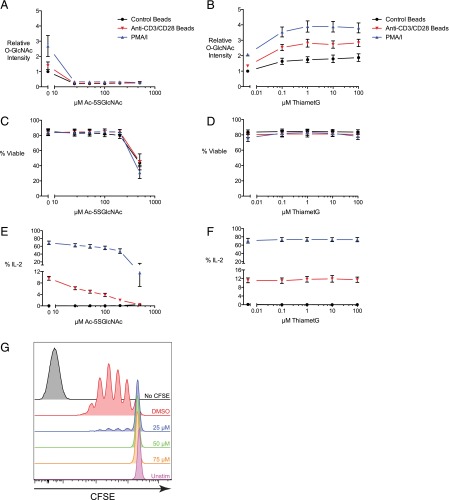
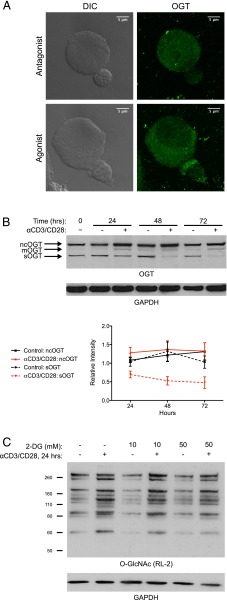
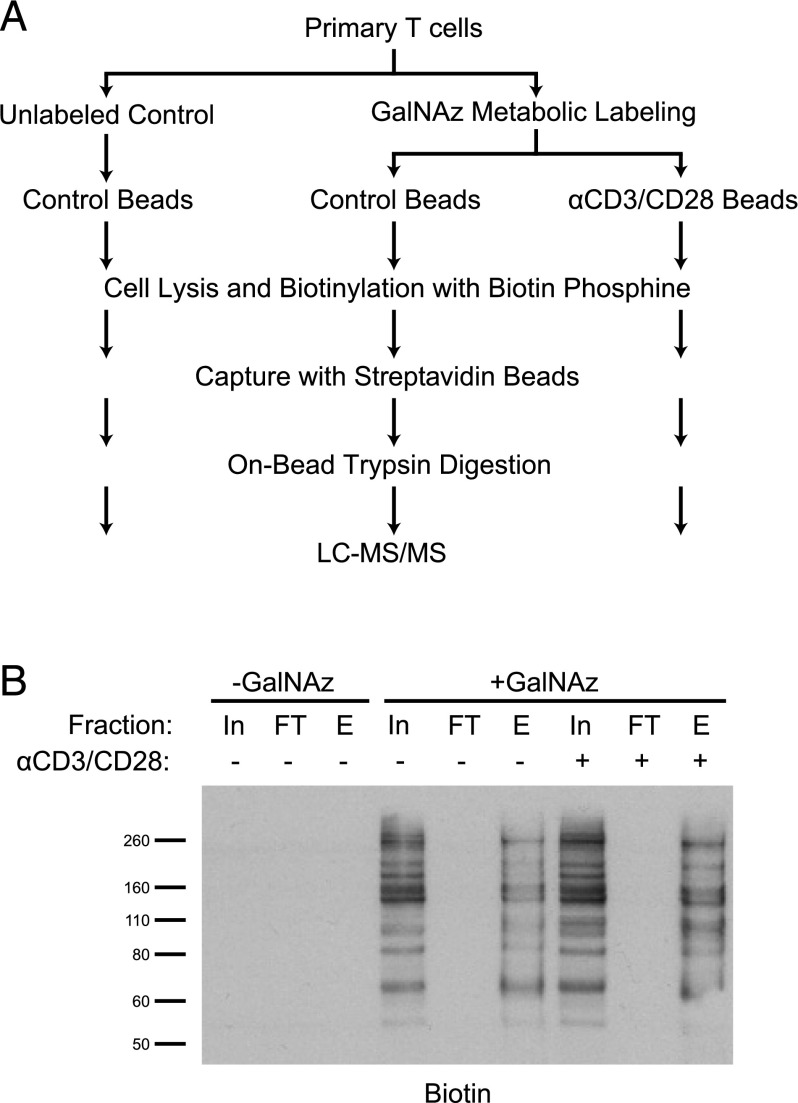
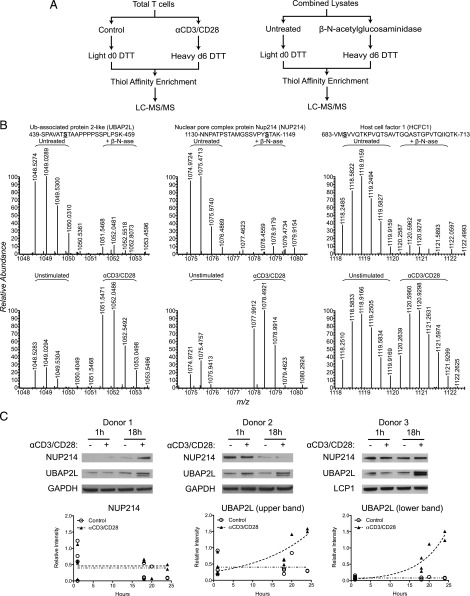
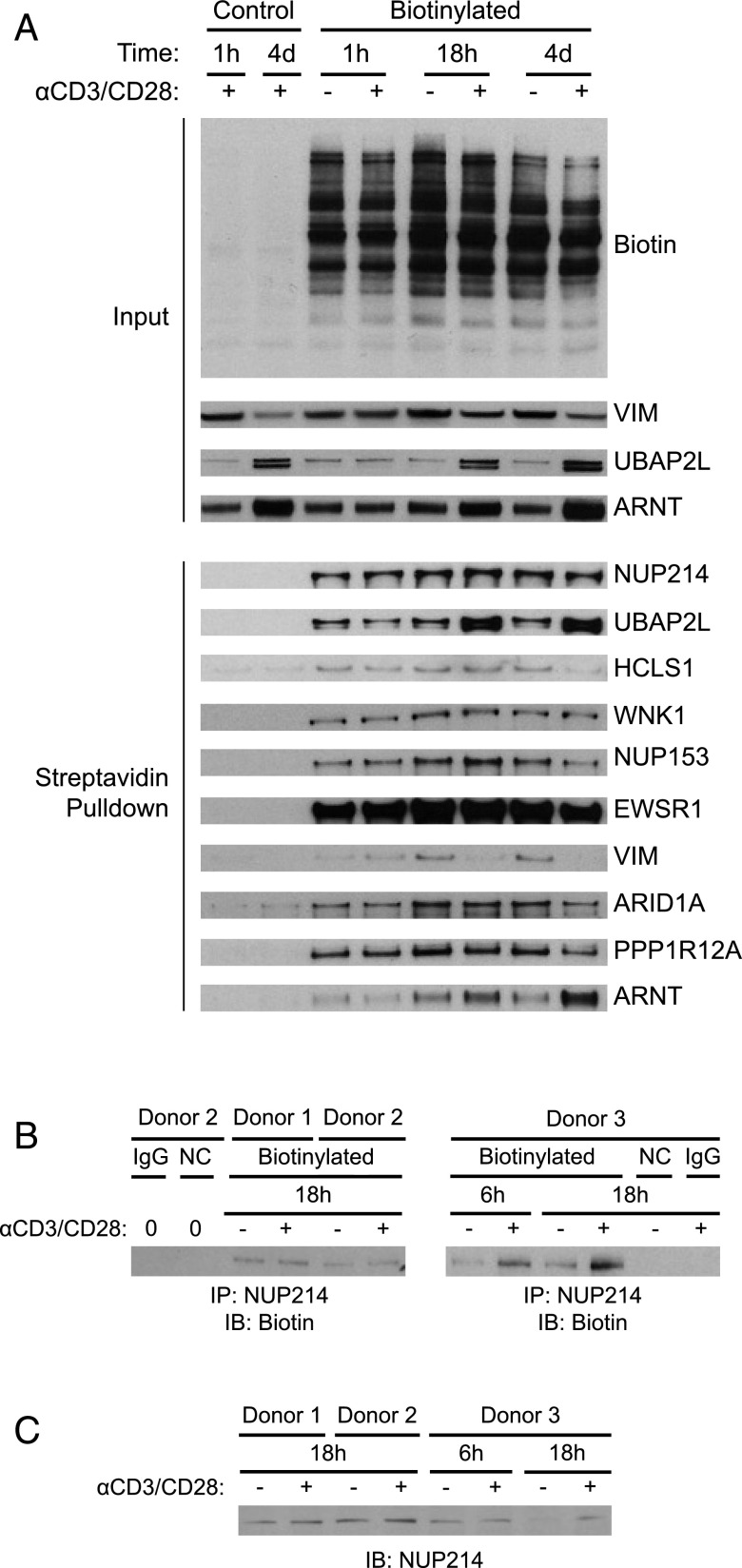
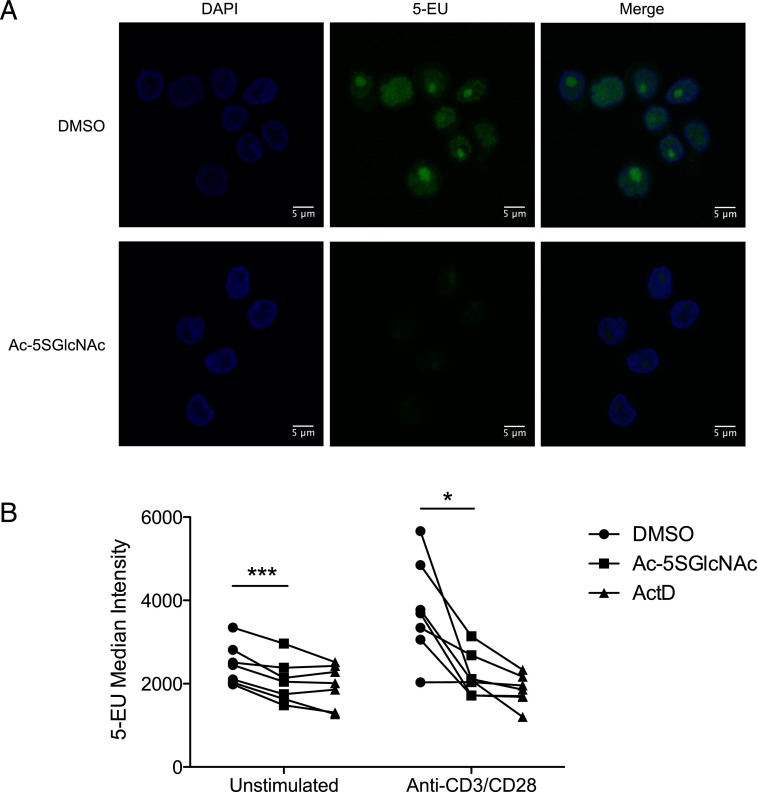
T cell activation in response to Ag is largely regulated by protein posttranslational modifications. Although phosphorylation has been extensively characterized in T cells, much less is known about the glycosylation of serine/threonine residues by O-linked N-acetylglucosamine (O-GlcNAc). Given that O-GlcNAc appears to regulate cell signaling pathways and protein activity similarly to phosphorylation, we performed a comprehensive analysis of O-GlcNAc during T cell activation to address the functional importance of this modification and to identify the modified proteins. Activation of T cells through the TCR resulted in a global elevation of O-GlcNAc levels and in the absence of O-GlcNAc, IL-2 production and proliferation were compromised. T cell activation also led to changes in the relative expression of O-GlcNAc transferase (OGT) isoforms and accumulation of OGT at the immunological synapse of murine T cells. Using a glycoproteomics approach, we identified >200 O-GlcNAc proteins in human T cells. Many of the identified proteins had a functional relationship to RNA metabolism, and consistent with a connection between O-GlcNAc and RNA, inhibition of OGT impaired nascent RNA synthesis upon T cell activation. Overall, our studies provide a global analysis of O-GlcNAc dynamics during T cell activation and the first characterization, to our knowledge, of the O-GlcNAc glycoproteome in human T cells.
Copyright © 2016 by The American Association of Immunologists, Inc.
Figures
References
-
- Acuto O., Di Bartolo V., Michel F. 2008. Tailoring T-cell receptor signals by proximal negative feedback mechanisms. Nat. Rev. Immunol. 8: 699–712. - PubMed
-
- Avni O., Lee D., Macian F., Szabo S. J., Glimcher L. H., Rao A. 2002. T(H) cell differentiation is accompanied by dynamic changes in histone acetylation of cytokine genes. Nat. Immunol. 3: 643–651. - PubMed
-
- Anandasabapathy N., Ford G. S., Bloom D., Holness C., Paragas V., Seroogy C., Skrenta H., Hollenhorst M., Fathman C. G., Soares L. 2003. GRAIL: an E3 ubiquitin ligase that inhibits cytokine gene transcription is expressed in anergic CD4+ T cells. Immunity 18: 535–547. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
