

Sexually dimorphic brain fatty acid composition in low and high fat diet-fed mice
- PMID: 27656405
- PMCID: PMC5021676
- DOI: 10.1016/j.molmet.2016.06.014
Sexually dimorphic brain fatty acid composition in low and high fat diet-fed mice
Abstract
Objective: In this study, we analyzed the fatty acid profile of brains and plasma from male and female mice fed chow or a western-style high fat diet (WD) for 16 weeks to determine if males and females process fatty acids differently. Based on the differences in fatty acids observed in vivo, we performed in vitro experiments on N43 hypothalamic neuronal cells to begin to elucidate how the fatty acid milieu may impact brain inflammation.
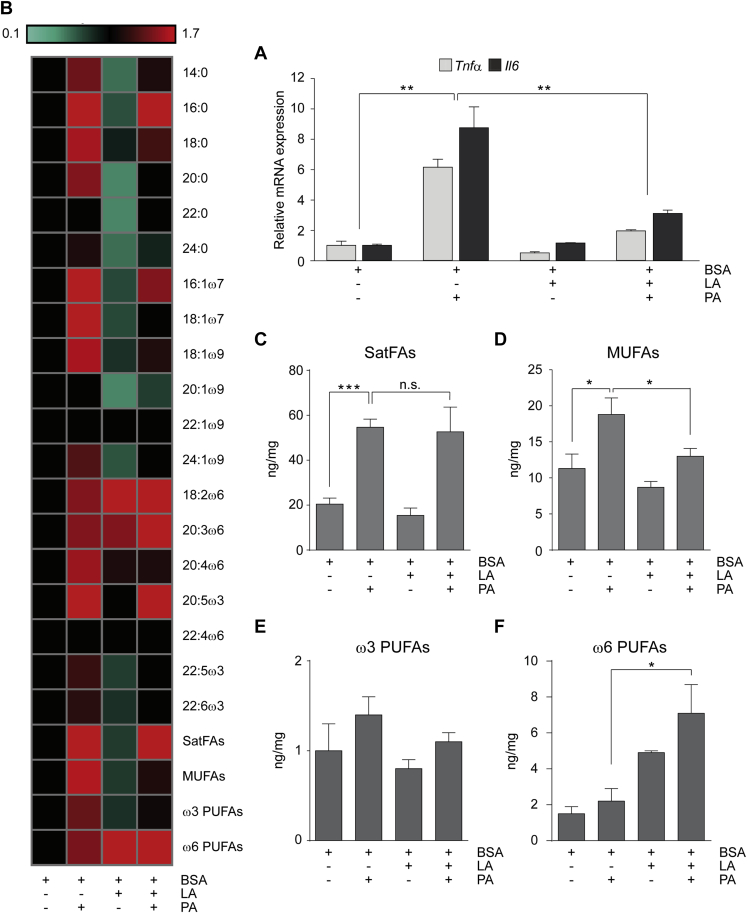
Methods: Using a comprehensive mass spectrometry fatty acid analysis, which includes a profile for 52 different fatty acid isomers, we assayed the plasma and brain fatty acid composition of age-matched male and female mice maintained on chow or a WD. Additionally, using the same techniques, we determined the fatty acid composition of N43 hypothalamic cells following exposure to palmitic and linoleic acid, alone or in combination.
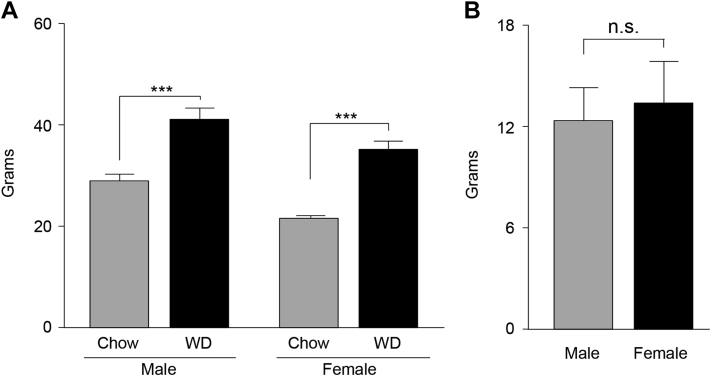
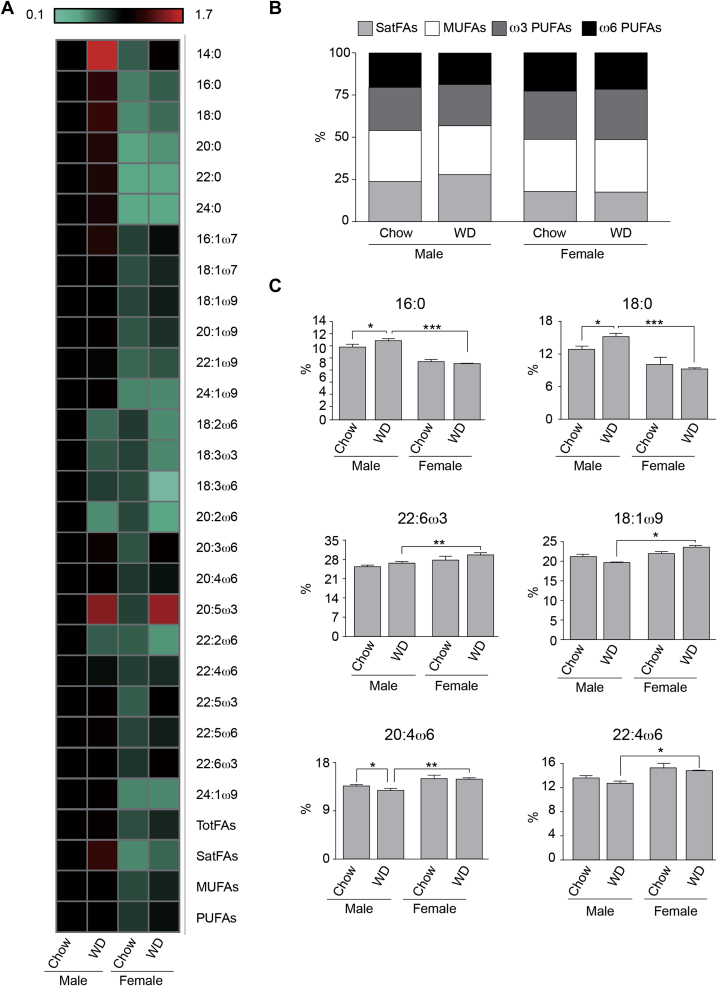
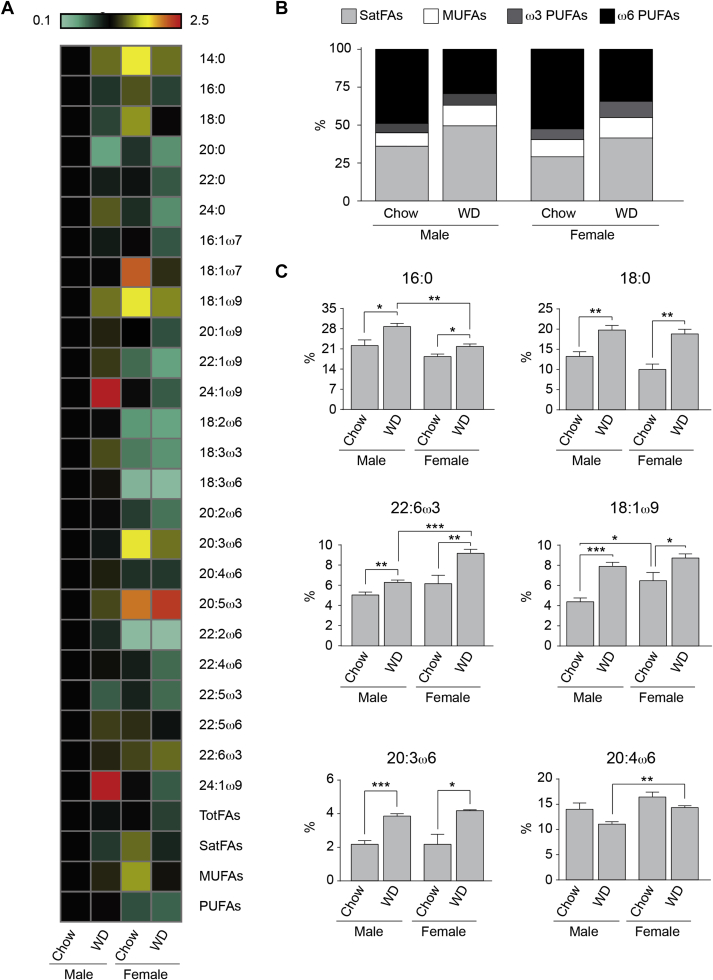
Results: Our data demonstrate there is a sexual dimorphism in brain fatty acid content both following the consumption of the chow diet, as well as the WD, with males having an increased percentage of saturated fatty acids and reductions in ω6-polyunsaturated fatty acids when compared to females. Interestingly, we did not observe a sexual dimorphism in fatty acid content in the plasma of the same mice. Furthermore, exposure of N43 cells to the ω6-PUFA linoleic acid, which is higher in female brains when compared to males, reduces palmitic acid-induced inflammation.
Conclusions: Our data suggest male and female brains, and not plasma, differ in their fatty acid profile. This is the first time, to our knowledge, lipidomic analyses has been used to directly test the hypothesis there is a sexual dimorphism in brain and plasma fatty acid composition following consumption of the chow diet, as well as following exposure to the WD.
Keywords: AA, arachidonic acid; ACC, acetyl-CoA carboxylase; B2m, beta-2 microglobulin; BBB, blood brain barrier; BSA, bovine serum albumin; C, Chow diet; CNS, central nervous system; Central nervous system; DHA, docosahexaenoic acid; F, female; FABP, fatty acid binding protein; FAS, fatty acid synthase; FAT/CD36, fatty acid transporter; FATP1, fatty acid transport protein 1; FAs, fatty acids; FFAs, free fatty acids; IL6, interleukin 6; LA, linoleic acid; Linoleic acid; M, male; MCD, malonyl-CoA decarboxylase; MSFD2a, membrane protein major facilitator super family domain containing 2a; MUFAs, monounsaturated fatty acids; N43; NF-κB, Nuclear Factor-κ Beta; Obesity; PA, palmitic acid; PUFAs, polyunsaturated fatty acids; Palmitic acid; SatFAs, saturated fatty acids; TFAs, total fatty acids; TNFα, Tumor Necrosis Factor α; UnsatFAs, unsaturated fatty acids; WD, western diet; WT, wild-type; Western diet; ω6-fatty acids.
Figures
References
-
- Carrie I., Clement M., de Javel D., Frances H., Bourre J.M. Specific phospholipid fatty acid composition of brain regions in mice. Effects of n-3 polyunsaturated fatty acid deficiency and phospholipid supplementation. Journal of Lipid Research. 2000;41(3):465–472. - PubMed
-
- Chang C.Y., Ke D.S., Chen J.Y. Essential fatty acids and human brain. Acta Neurologica Taiwanica. 2009;18(4):231–241. - PubMed
-
- Chen C.T., Green J.T., Orr S.K., Bazinet R.P. Regulation of brain polyunsaturated fatty acid uptake and turnover. Prostaglandins, Leukotrienes and Essential Fatty Acids. 2008;79(3–5):85–91. - PubMed
-
- Bazinet R.P., Laye S. Polyunsaturated fatty acids and their metabolites in brain function and disease. Nature Reviews Neuroscience. 2014;15(12):771–785. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
