

Arms Race between Enveloped Viruses and the Host ERAD Machinery
- PMID: 27657106
- PMCID: PMC5035969
- DOI: 10.3390/v8090255
Arms Race between Enveloped Viruses and the Host ERAD Machinery
Abstract
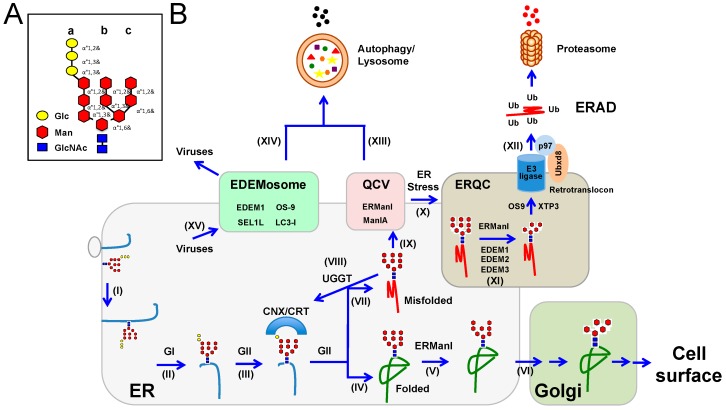
Enveloped viruses represent a significant category of pathogens that cause serious diseases in animals. These viruses express envelope glycoproteins that are singularly important during the infection of host cells by mediating fusion between the viral envelope and host cell membranes. Despite low homology at protein levels, three classes of viral fusion proteins have, as of yet, been identified based on structural similarities. Their incorporation into viral particles is dependent upon their proper sub-cellular localization after being expressed and folded properly in the endoplasmic reticulum (ER). However, viral protein expression can cause stress in the ER, and host cells respond to alleviate the ER stress in the form of the unfolded protein response (UPR); the effects of which have been observed to potentiate or inhibit viral infection. One important arm of UPR is to elevate the capacity of the ER-associated protein degradation (ERAD) pathway, which is comprised of host quality control machinery that ensures proper protein folding. In this review, we provide relevant details regarding viral envelope glycoproteins, UPR, ERAD, and their interactions in host cells.
Keywords: ER stress; ERAD; UPR; endoplasmic reticulum-associated degradation; enveloped viruses; unfolded protein response; viral glycoproteins.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Viral mediated tethering to SEL1L facilitates ER-associated degradation of IRE1.J Virol. 2021 Mar 25;95(8):e01990-20. doi: 10.1128/JVI.01990-20. Epub 2021 Jan 20. J Virol. 2021. PMID: 33472927 Free PMC article.
-
Mif1: a missing link between the unfolded protein response pathway and ER-associated protein degradation?Curr Protein Pept Sci. 2001 Jun;2(2):169-90. doi: 10.2174/1389203013381189. Curr Protein Pept Sci. 2001. PMID: 12370023 Review.
-
The ER quality control and ER associated degradation machineries are vital for viral pathogenesis.Front Plant Sci. 2014 Mar 11;5:66. doi: 10.3389/fpls.2014.00066. eCollection 2014. Front Plant Sci. 2014. PMID: 24653727 Free PMC article. Review.
-
Herpes Simplex Virus 1 UL41 Protein Suppresses the IRE1/XBP1 Signal Pathway of the Unfolded Protein Response via Its RNase Activity.J Virol. 2017 Jan 31;91(4):e02056-16. doi: 10.1128/JVI.02056-16. Print 2017 Feb 15. J Virol. 2017. PMID: 27928013 Free PMC article.
-
Immature Core protein of hepatitis C virus induces an unfolded protein response through inhibition of ERAD-L in a yeast model system.Genes Cells. 2017 Feb;22(2):160-173. doi: 10.1111/gtc.12464. Epub 2017 Jan 18. Genes Cells. 2017. PMID: 28097745
Cited by
-
SYNJ2BP Improves the Production of Lentiviral Envelope Protein by Facilitating the Formation of Mitochondrion-Associated Endoplasmic Reticulum Membrane.J Virol. 2022 Oct 26;96(20):e0054922. doi: 10.1128/jvi.00549-22. Epub 2022 Oct 5. J Virol. 2022. PMID: 36197105 Free PMC article.
-
Organelle dynamics and viral infections: at cross roads.Microbes Infect. 2019 Jan-Feb;21(1):20-32. doi: 10.1016/j.micinf.2018.06.002. Epub 2018 Jun 25. Microbes Infect. 2019. PMID: 29953921 Free PMC article. Review.
-
Antiviral strategies targeting host factors and mechanisms obliging +ssRNA viral pathogens.Bioorg Med Chem. 2021 Sep 15;46:116356. doi: 10.1016/j.bmc.2021.116356. Epub 2021 Aug 8. Bioorg Med Chem. 2021. PMID: 34416512 Free PMC article. Review.
-
The H protein of attenuated canine distemper virus is degraded via endoplasmic reticulum-associated protein degradation.Front Vet Sci. 2023 Jul 6;10:1214318. doi: 10.3389/fvets.2023.1214318. eCollection 2023. Front Vet Sci. 2023. PMID: 37483299 Free PMC article.
-
Non-lysine ubiquitylation: Doing things differently.Front Mol Biosci. 2022 Sep 19;9:1008175. doi: 10.3389/fmolb.2022.1008175. eCollection 2022. Front Mol Biosci. 2022. PMID: 36200073 Free PMC article. Review.
References
-
- Knipe D.M., Howley P.M. Fields Virology. 6th ed. Wolters Kluwer/Lippincott Williams & Wilkins Health; Philadelphia, PA, USA: 2013.
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
