

Optogenetic activation of melanin-concentrating hormone neurons increases non-rapid eye movement and rapid eye movement sleep during the night in rats
- PMID: 27657541
- PMCID: PMC5118149
- DOI: 10.1111/ejn.13410
Optogenetic activation of melanin-concentrating hormone neurons increases non-rapid eye movement and rapid eye movement sleep during the night in rats
Abstract
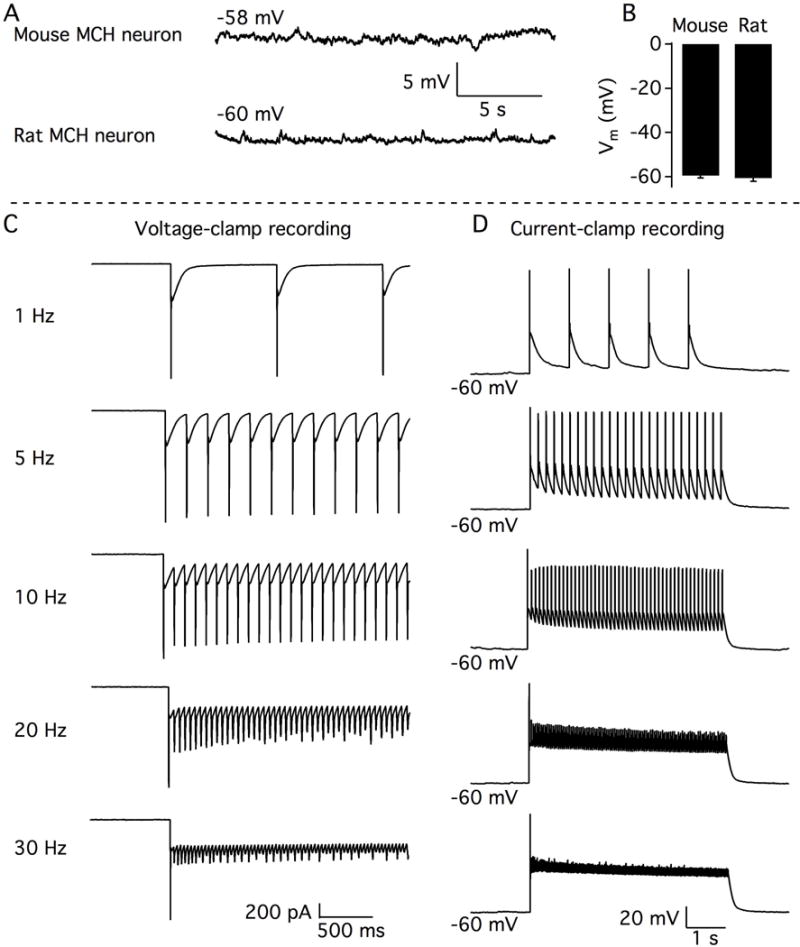
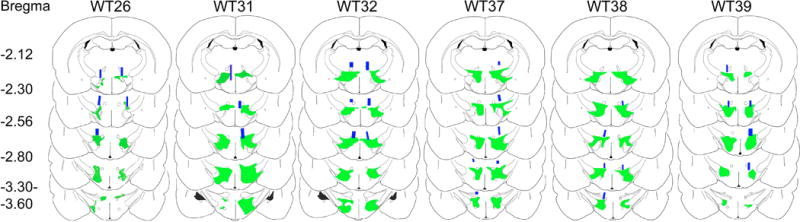
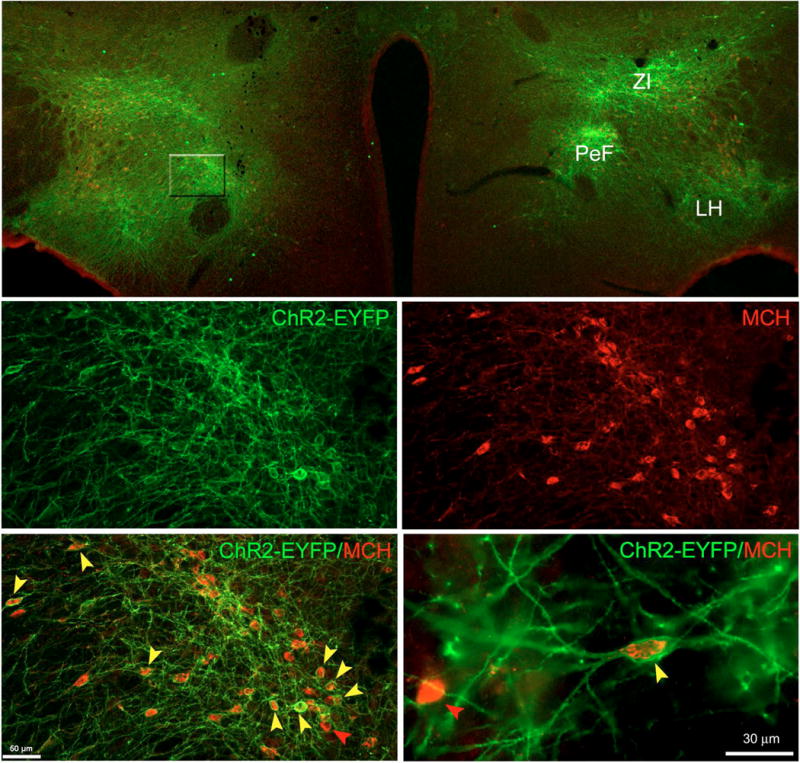
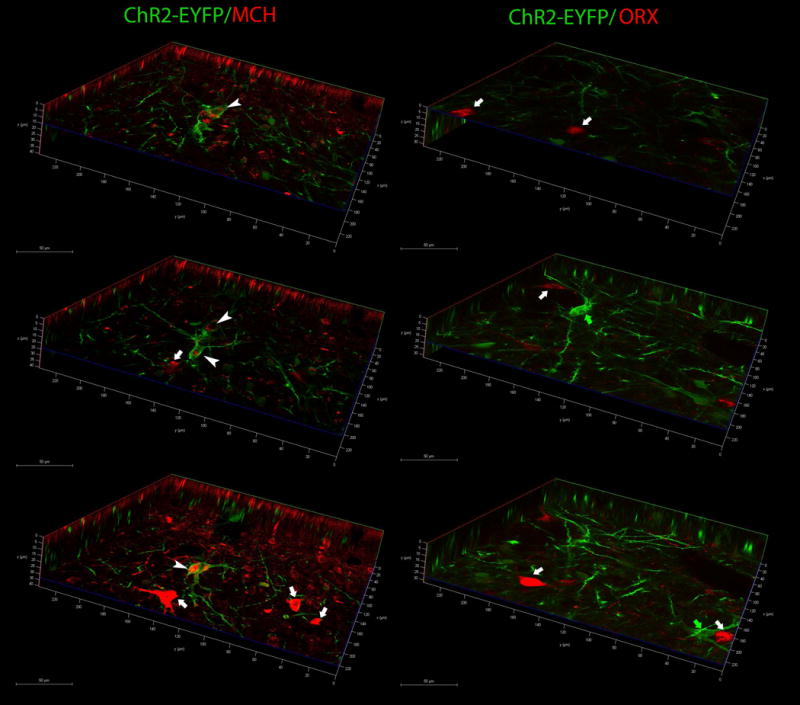
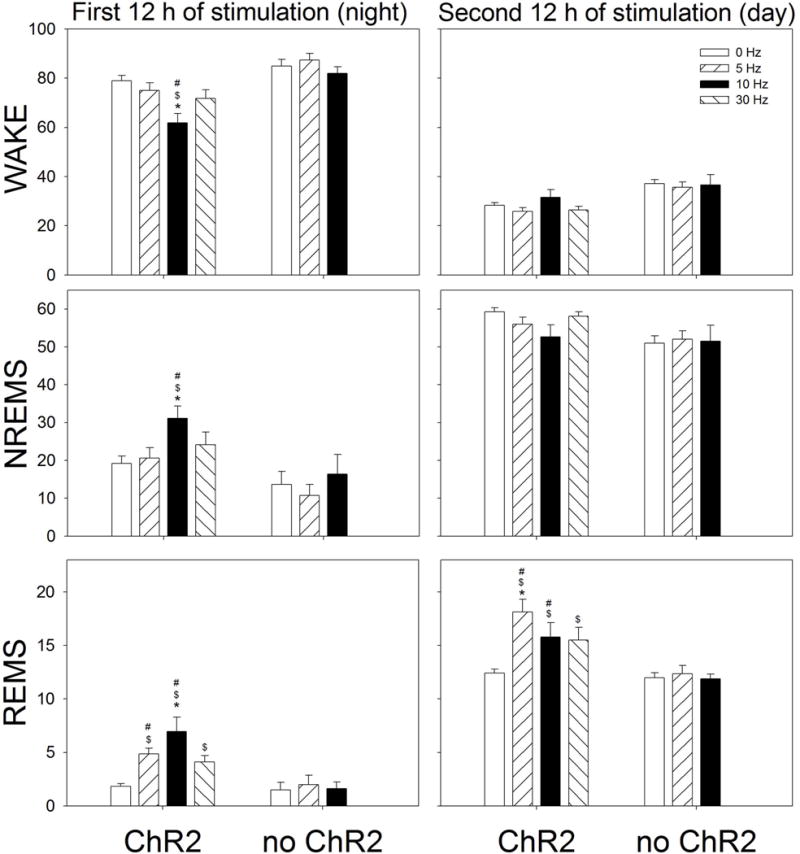
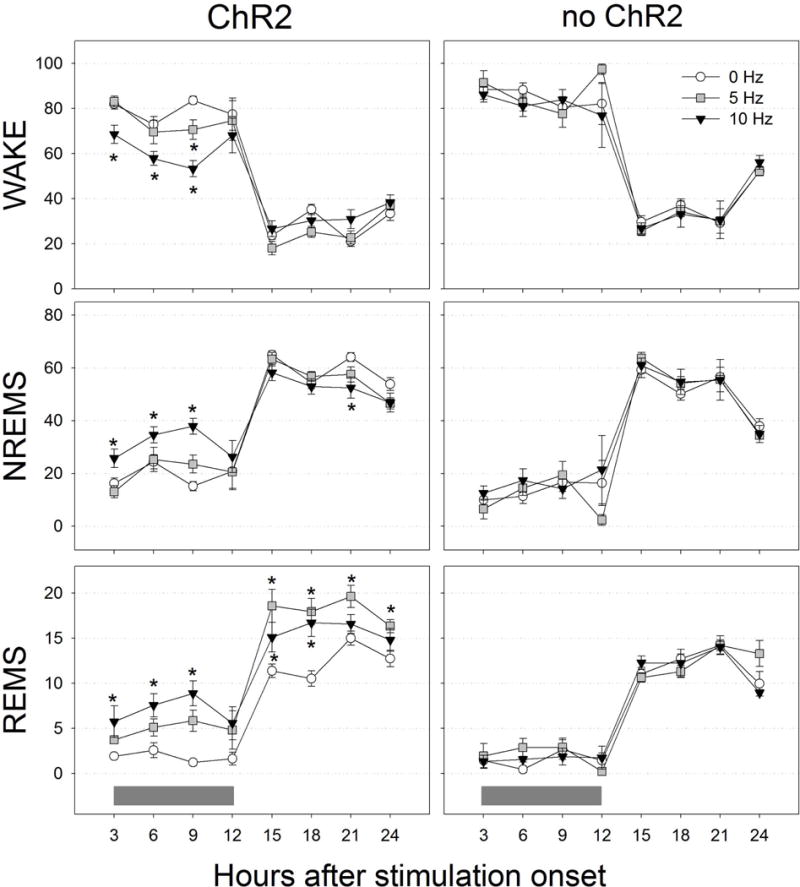
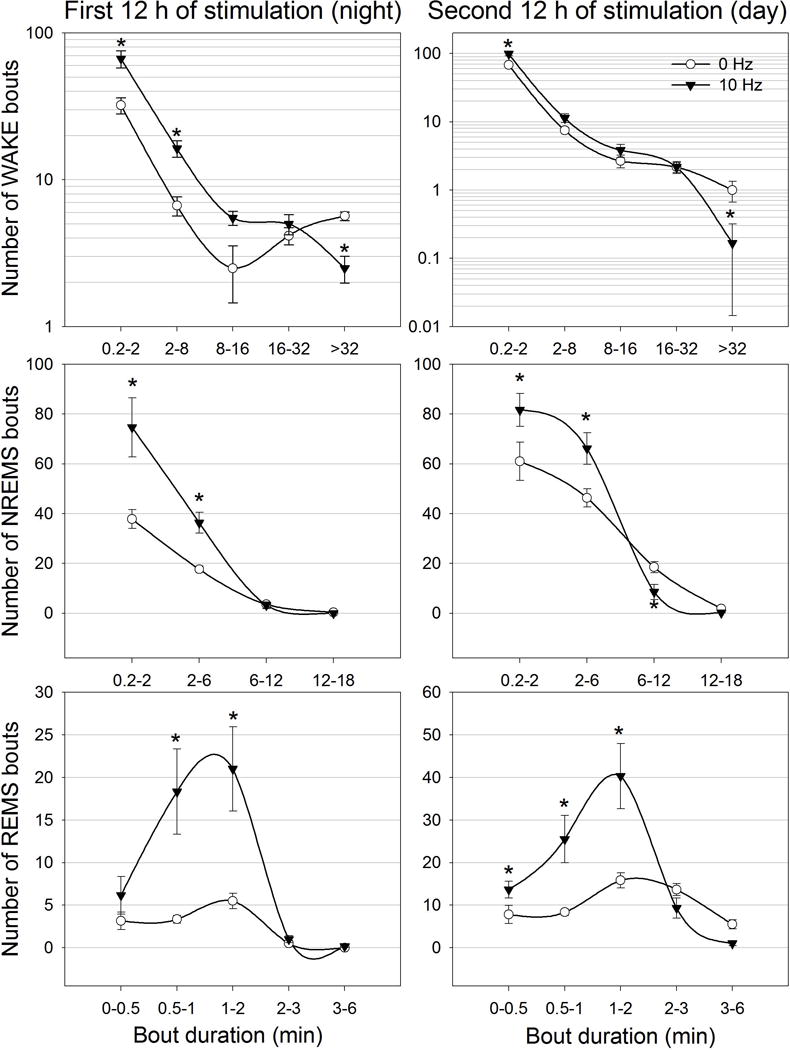
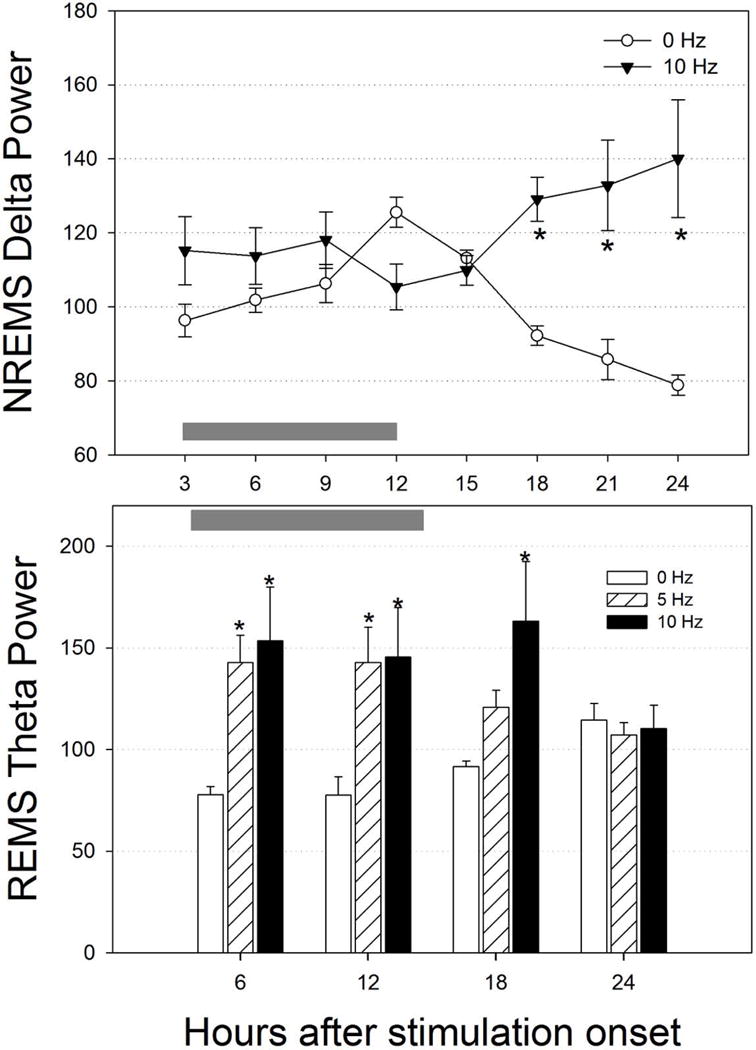
Neurons containing melanin-concentrating hormone (MCH) are located in the hypothalamus. In mice, optogenetic activation of the MCH neurons induces both non-rapid eye movement (NREM) and rapid eye movement (REM) sleep at night, the normal wake-active period for nocturnal rodents [R. R. Konadhode et al. (2013) J. Neurosci., 33, 10257-10263]. Here we selectively activate these neurons in rats to test the validity of the sleep network hypothesis in another species. Channelrhodopsin-2 (ChR2) driven by the MCH promoter was selectively expressed by MCH neurons after injection of rAAV-MCHp-ChR2-EYFP into the hypothalamus of Long-Evans rats. An in vitro study confirmed that the optogenetic activation of MCH neurons faithfully triggered action potentials. In the second study, in Long-Evans rats, rAAV-MCH-ChR2, or the control vector, rAAV-MCH-EYFP, were delivered into the hypothalamus. Three weeks later, baseline sleep was recorded for 48 h without optogenetic stimulation (0 Hz). Subsequently, at the start of the lights-off cycle, the MCH neurons were stimulated at 5, 10, or 30 Hz (1 mW at tip; 1 min on - 4 min off) for 24 h. Sleep was recorded during the 24-h stimulation period. Optogenetic activation of MCH neurons increased both REM and NREM sleep at night, whereas during the day cycle, only REM sleep was increased. Delta power, an indicator of sleep intensity, was also increased. In control rats without ChR2, optogenetic stimulation did not increase sleep or delta power. These results lend further support to the view that sleep-active MCH neurons contribute to drive sleep in mammals.
Keywords: channelrhodopsin-2; melanin-concentrating hormone; rapid eye movement sleep; rat; recombinant adeno-associated virus.
© 2016 Federation of European Neuroscience Societies and John Wiley & Sons Ltd.
Conflict of interest statement
No conflicts of interest, financial or otherwise are declared by the author(s).
Figures
References
-
- Ahnaou A, Dautzenberg FM, Huysmans H, Steckler T, Drinkenburg WH. Contribution of melanin-concentrating hormone (MCH1) receptor to thermoregulation and sleep stabilization: evidence from MCH1 (−/−) mice. Behavioural brain research. 2011;218:42–50. - PubMed
-
- Ahnaou A, Drinkenburg WH, Bouwknecht JA, Alcazar J, Steckler T, Dautzenberg FM. Blocking melanin-concentrating hormone MCH1 receptor affects rat sleep-wake architecture. European journal of pharmacology. 2008;579:177–188. - PubMed
-
- Apergis-Schoute J, Iordanidou P, Faure C, Jego S, Schone C, Aitta-Aho T, Adamantidis A, Burdakov D. Optogenetic evidence for inhibitory signaling from orexin to MCH neurons via local microcircuits. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2015;35:5435–5441. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
